

The WH2 Domain and Actin Nucleation: Necessary but Insufficient
- PMID: 27068179
- PMCID: PMC4884163
- DOI: 10.1016/j.tibs.2016.03.004
The WH2 Domain and Actin Nucleation: Necessary but Insufficient
Abstract
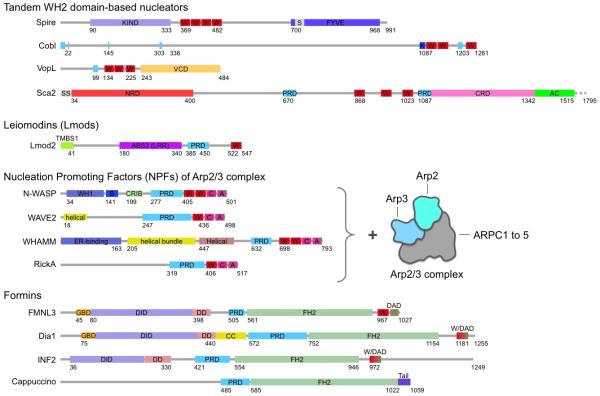
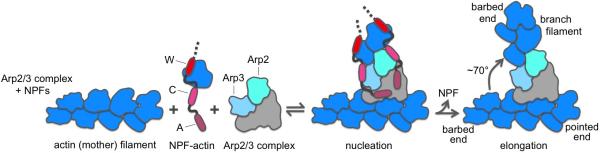
Two types of sequences, proline-rich domains (PRDs) and the WASP-homology 2 (WH2) domain, are found in most actin filament nucleation and elongation factors discovered thus far. PRDs serve as a platform for protein-protein interactions, often mediating the binding of profilin-actin. The WH2 domain is an abundant actin monomer-binding motif comprising ∼17 amino acids. It frequently occurs in tandem repeats, and functions in nucleation by recruiting actin subunits to form the polymerization nucleus. It is found in Spire, Cordon Bleu (Cobl), Leiomodin (Lmod), Arp2/3 complex activators (WASP, WHAMM, WAVE, etc.), the bacterial nucleators VopL/VopF and Sca2, and some formins. Yet, it is argued here that the WH2 domain plays only an auxiliary role in nucleation, always synergizing with other domains or proteins for this activity.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures




References
-
- Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. - PubMed
-
- Dietrich S, et al. Structural and functional insights into the Spir/formin actin nucleator complex. Biol Chem. 2013;394:1649–1660. - PubMed
-
- Carlier MF, et al. Control of actin assembly by the WH2 domains and their multifunctional tandem repeats in Spire and Cordon-Bleu. Int Rev Cell Mol Biol. 2011;290:55–85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
