

BET Bromodomain Inhibition Releases the Mediator Complex from Select cis-Regulatory Elements
- PMID: 27068464
- PMCID: PMC4838499
- DOI: 10.1016/j.celrep.2016.03.054
BET Bromodomain Inhibition Releases the Mediator Complex from Select cis-Regulatory Elements
Abstract
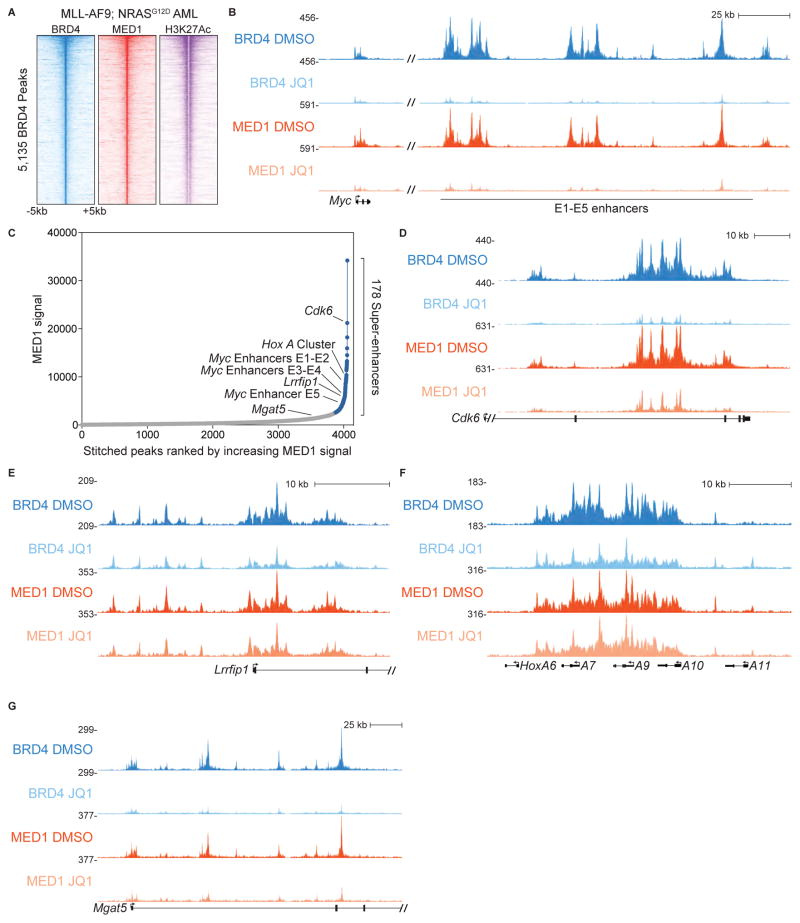
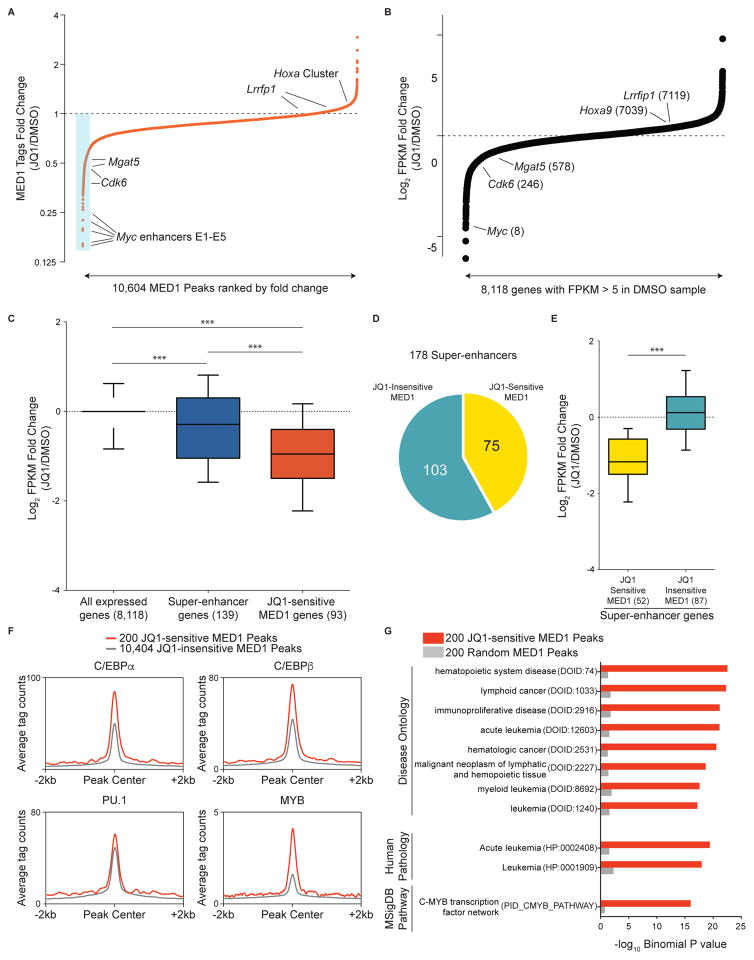
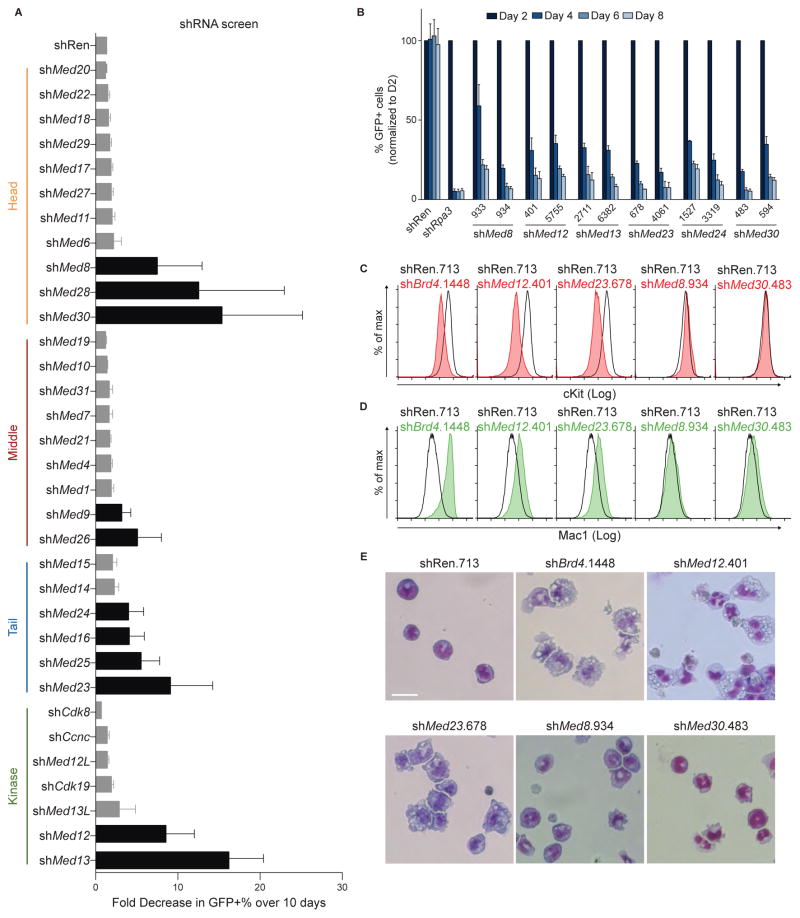
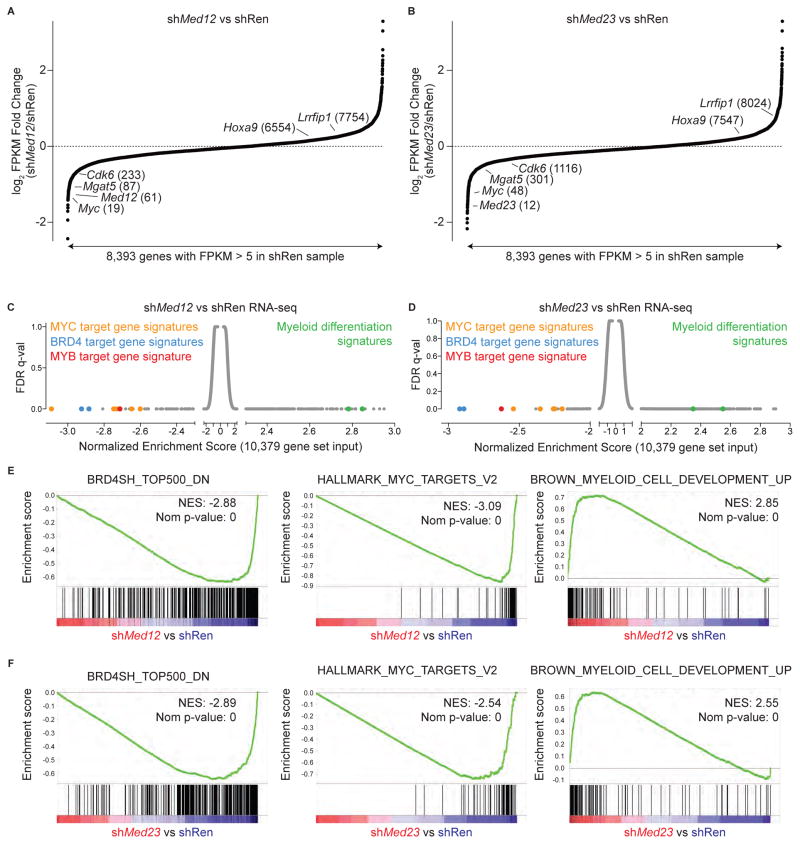
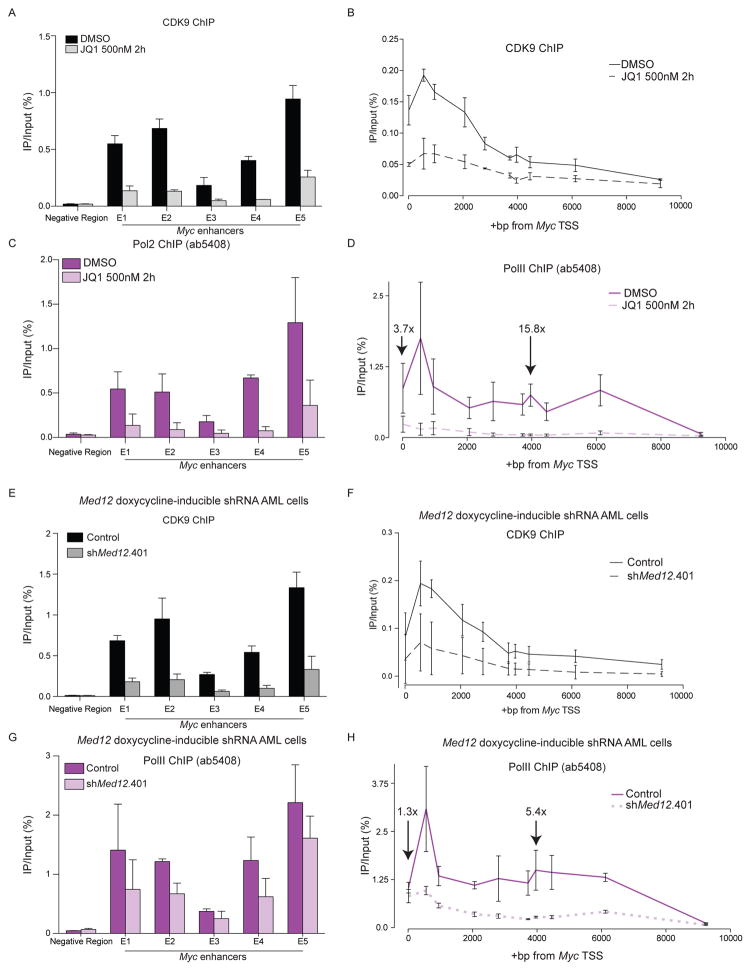
The bromodomain and extraterminal (BET) protein BRD4 can physically interact with the Mediator complex, but the relevance of this association to the therapeutic effects of BET inhibitors in cancer is unclear. Here, we show that BET inhibition causes a rapid release of Mediator from a subset of cis-regulatory elements in the genome of acute myeloid leukemia (AML) cells. These sites of Mediator eviction were highly correlated with transcriptional suppression of neighboring genes, which are enriched for targets of the transcription factor MYB and for functions related to leukemogenesis. A shRNA screen of Mediator in AML cells identified the MED12, MED13, MED23, and MED24 subunits as performing a similar regulatory function to BRD4 in this context, including a shared role in sustaining a block in myeloid maturation. These findings suggest that the interaction between BRD4 and Mediator has functional importance for gene-specific transcriptional activation and for AML maintenance.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Di Micco R, Fontanals-Cirera B, Low V, Ntziachristos P, Yuen SK, Lovell CD, Dolgalev I, Yonekubo Y, Zhang G, Rusinova E, et al. Control of embryonic stem cell identity by BRD4-dependent transcriptional elongation of super-enhancer-associated pluripotency genes. Cell reports. 2014;9:234–247. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
