

Naturally occurring mitochondrial-derived peptides are age-dependent regulators of apoptosis, insulin sensitivity, and inflammatory markers
- PMID: 27070352
- PMCID: PMC4925829
- DOI: 10.18632/aging.100943
Naturally occurring mitochondrial-derived peptides are age-dependent regulators of apoptosis, insulin sensitivity, and inflammatory markers
Abstract
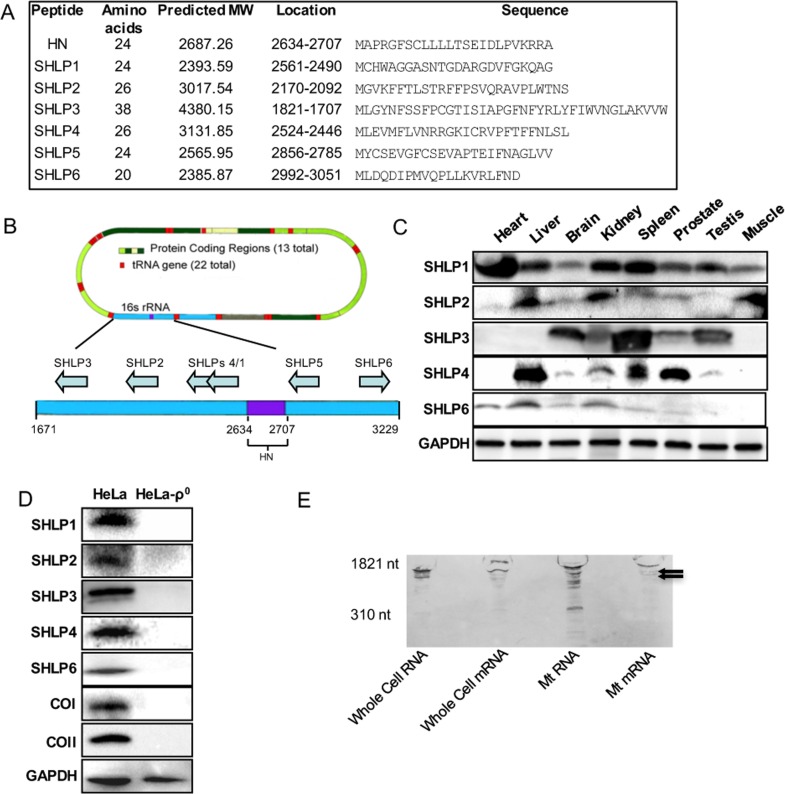
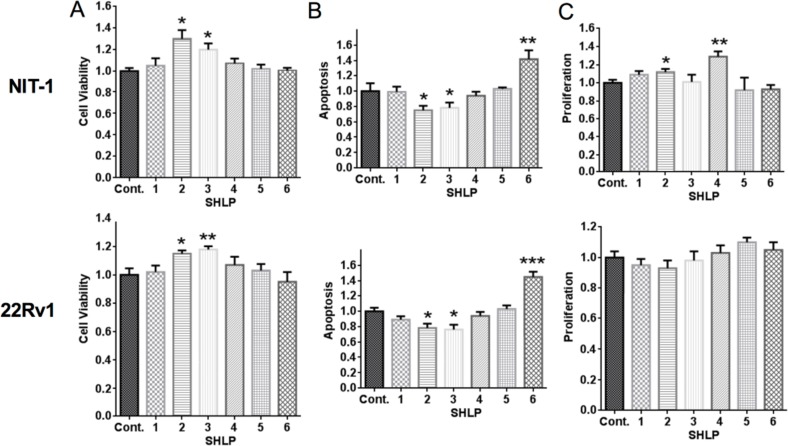
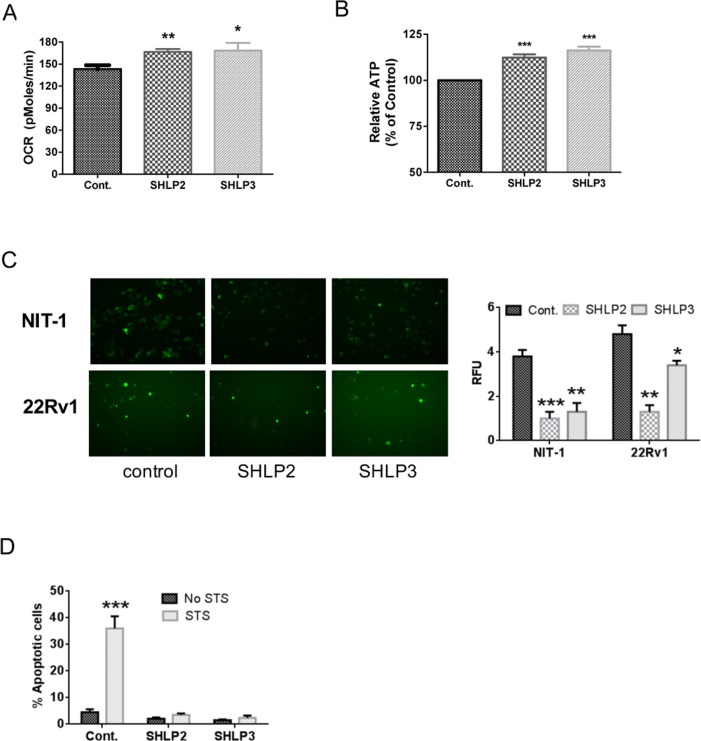
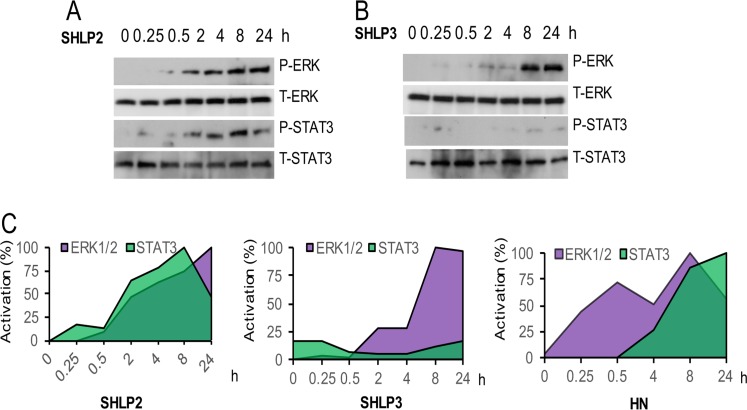
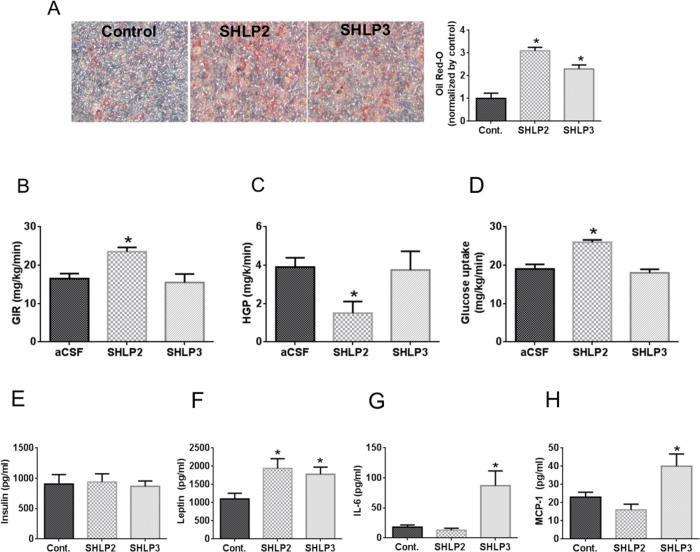
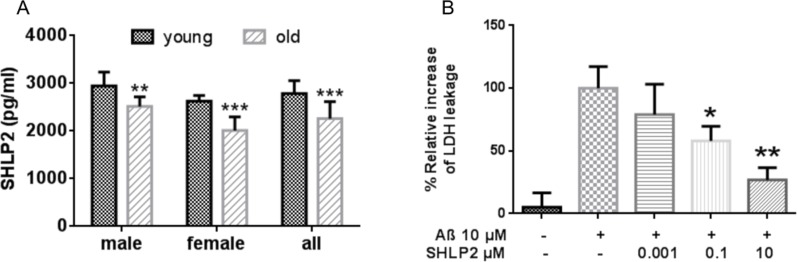
Mitochondria are key players in aging and in the pathogenesis of age-related diseases. Recent mitochondrial transcriptome analyses revealed the existence of multiple small mRNAs transcribed from mitochondrial DNA (mtDNA). Humanin (HN), a peptide encoded in the mtDNA 16S ribosomal RNA region, is a neuroprotective factor. An in silico search revealed six additional peptides in the same region of mtDNA as humanin; we named these peptides small humanin-like peptides (SHLPs). We identified the functional roles for these peptides and the potential mechanisms of action. The SHLPs differed in their ability to regulate cell viability in vitro. We focused on SHLP2 and SHLP3 because they shared similar protective effects with HN. Specifically, they significantly reduced apoptosis and the generation of reactive oxygen species, and improved mitochondrial metabolism in vitro. SHLP2 and SHLP3 also enhanced 3T3-L1 pre-adipocyte differentiation. Systemic hyperinsulinemic-euglycemic clamp studies showed that intracerebrally infused SHLP2 increased glucose uptake and suppressed hepatic glucose production, suggesting that it functions as an insulin sensitizer both peripherally and centrally. Similar to HN, the levels of circulating SHLP2 were found to decrease with age. These results suggest that mitochondria play critical roles in metabolism and survival through the synthesis of mitochondrial peptides, and provide new insights into mitochondrial biology with relevance to aging and human biology.
Keywords: SHLP; aging; humanin; mitochondria; small ORFs.
Conflict of interest statement
LC, CL, NB, and PC are consultants and stockholders of CohBar Inc.
Figures
References
-
- Hashimoto Y, Niikura T, Tajima H, Yasukawa T, Sudo H, Ito Y, Kita Y, Kawasumi M, Kouyama K, Doyu M, Sobue G, Koide T, Tsuji S, et al. A rescue factor abolishing neuronal cell death by a wide spectrum of familial Alzheimer's disease genes and Abeta. Proc Natl Acad Sci U S A. 2001;98:6336–6341. - PMC - PubMed
-
- Guo B, Zhai D, Cabezas E, Welsh K, Nouraini S, Satterthwait AC, Reed JC. Humanin peptide suppresses apoptosis by interfering with Bax activation. Nature. 2003;423:456–461. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
