

Anti-Inflammatory and Cytoprotective Effects of TMC-256C1 from Marine-Derived Fungus Aspergillus sp. SF-6354 via up-Regulation of Heme Oxygenase-1 in Murine Hippocampal and Microglial Cell Lines
- PMID: 27070586
- PMCID: PMC4848985
- DOI: 10.3390/ijms17040529
Anti-Inflammatory and Cytoprotective Effects of TMC-256C1 from Marine-Derived Fungus Aspergillus sp. SF-6354 via up-Regulation of Heme Oxygenase-1 in Murine Hippocampal and Microglial Cell Lines
Abstract
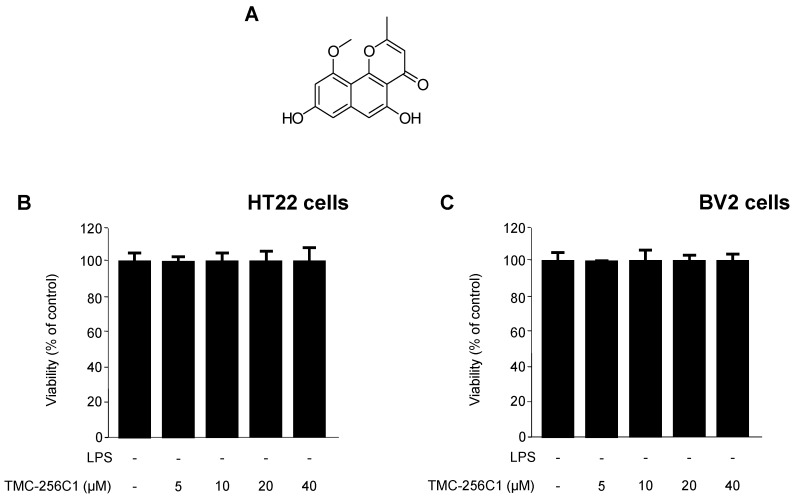
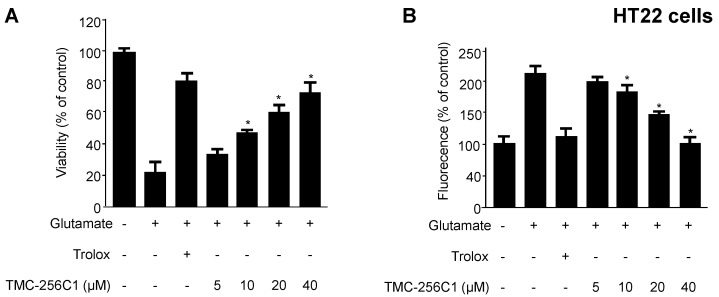
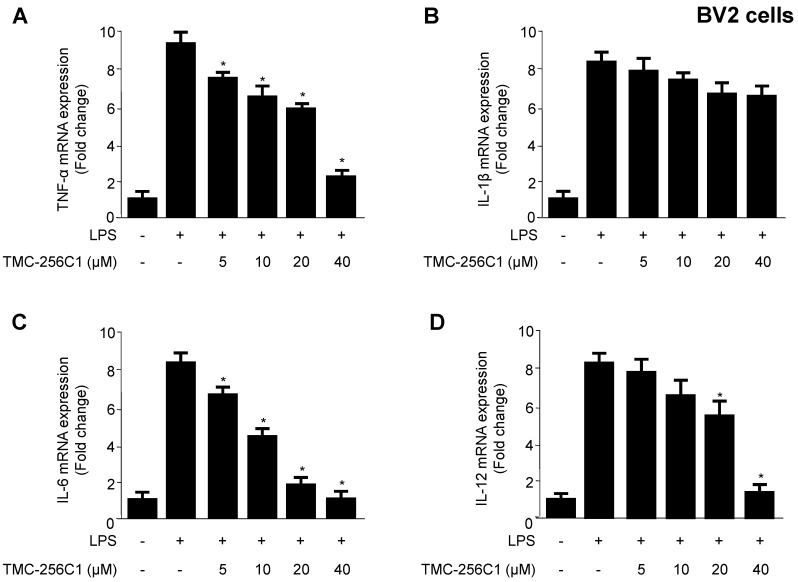
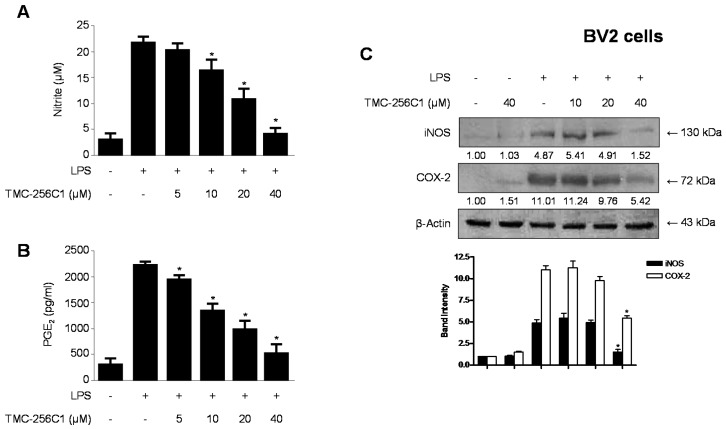
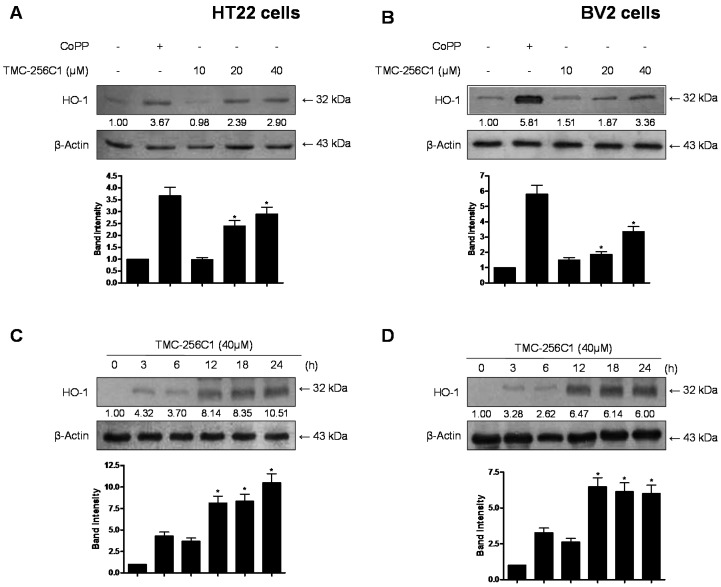
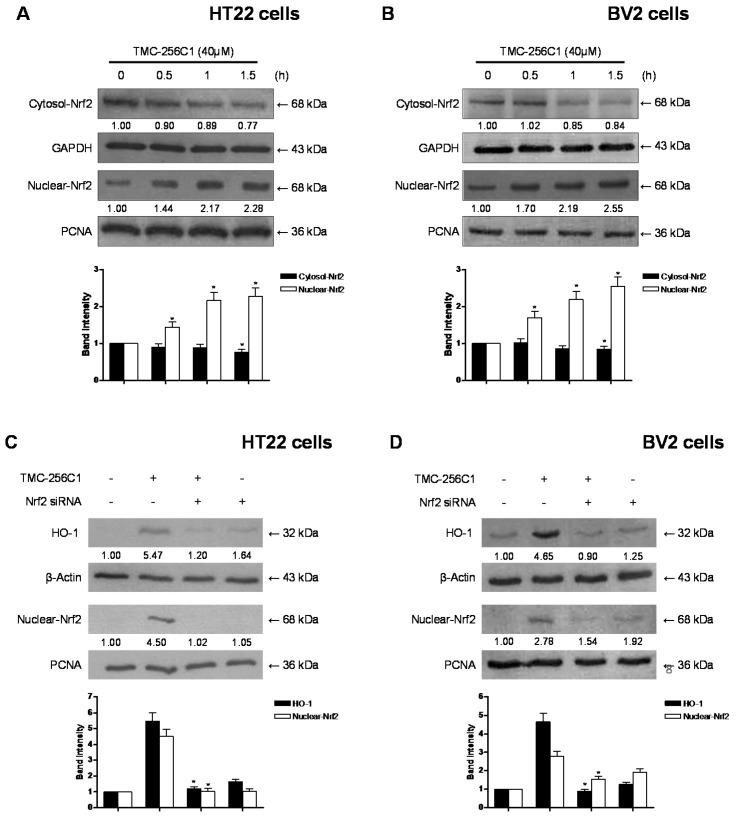
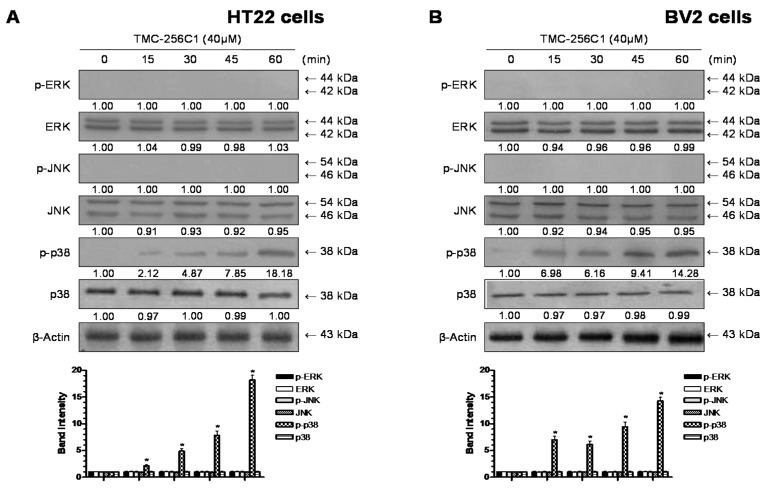
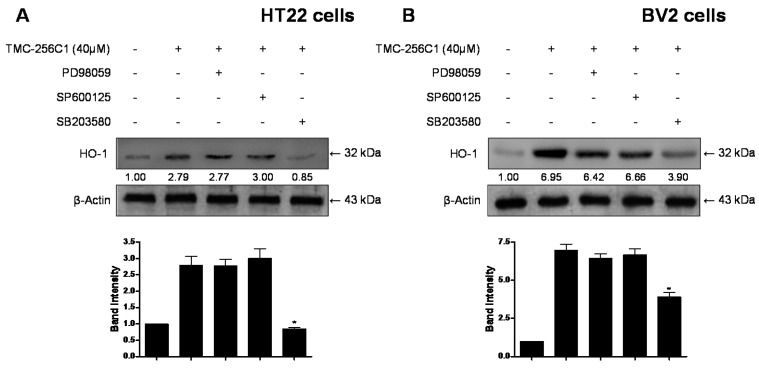
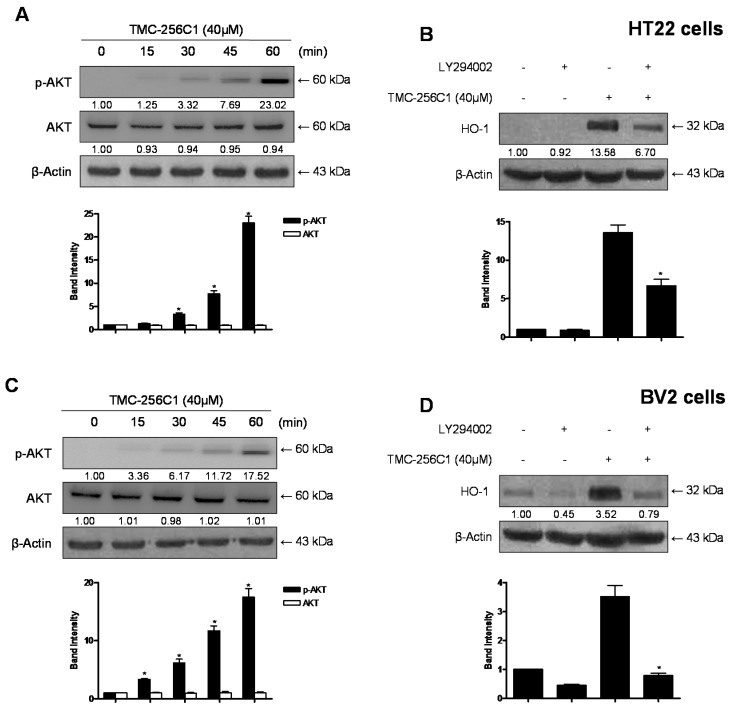
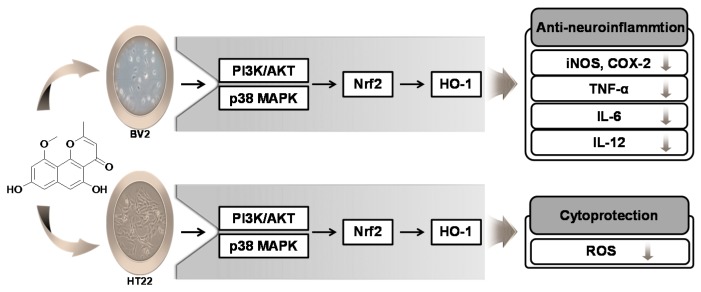
In the course of searching for bioactive secondary metabolites from marine fungi, TMC-256C1 was isolated from an ethyl acetate extract of the marine-derived fungus Aspergillus sp. SF6354. TMC-256C1 displayed anti-neuroinflammatory effect in BV2 microglial cells induced by lipopolysaccharides (LPS) as well as neuroprotective effect against glutamate-stimulated neurotoxicity in mouse hippocampal HT22 cells. TMC-256C1 was shown to develop a cellular resistance to oxidative damage caused by glutamate-induced cytotoxicity and reactive oxygen species (ROS) generation in HT22 cells, and suppress the inflammation process in LPS-stimulated BV2 cells. Furthermore, the neuroprotective and anti-neuroinflammatory activities of TMC-256C1 were associated with upregulated expression of heme oxygenase (HO)-1 and nuclear translocation of nuclear factor-E2-related factor 2 (Nrf2) in HT22 and BV2 cells. We also found that TMC-256C1 activated p38 mitogen-activated protein kinases (MAPK) and phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathways in HT22 and BV2 cells. These results demonstrated that TMC-256C1 activates HO-1 protein expression, probably by increasing nuclear Nrf2 levels via the activation of the p38 MAPK and PI3K/Akt pathways.
Keywords: Aspergillus; TMC-256C1; anti-neuroinflammatory effect; heme oxygenase-1 (HO-1); marine fungus; neuroprotective effect.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
