

Heterozygous colon cancer-associated mutations of SAMHD1 have functional significance
- PMID: 27071091
- PMCID: PMC4855590
- DOI: 10.1073/pnas.1519128113
Heterozygous colon cancer-associated mutations of SAMHD1 have functional significance
Erratum in
-
Correction for Rentoft et al., Heterozygous colon cancer-associated mutations of SAMHD1 have functional significance.Proc Natl Acad Sci U S A. 2019 Mar 5;116(10):4744. doi: 10.1073/pnas.1902081116. Epub 2019 Feb 25. Proc Natl Acad Sci U S A. 2019. PMID: 30804184 Free PMC article. No abstract available.
Abstract
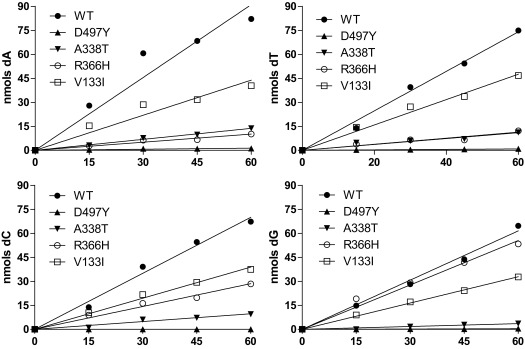
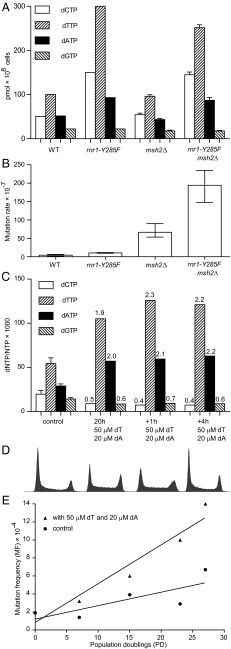
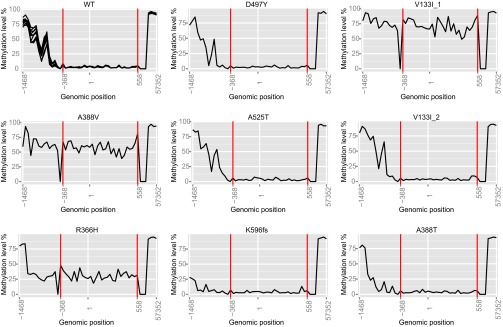
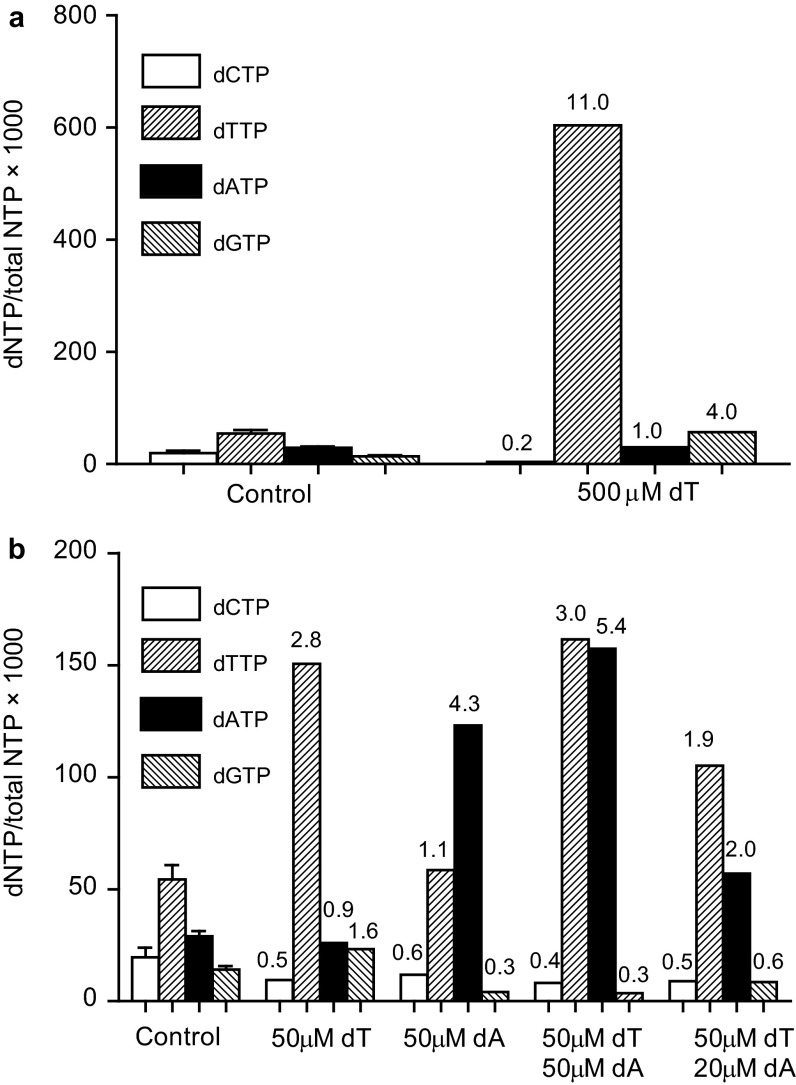
Even small variations in dNTP concentrations decrease DNA replication fidelity, and this observation prompted us to analyze genomic cancer data for mutations in enzymes involved in dNTP metabolism. We found that sterile alpha motif and histidine-aspartate domain-containing protein 1 (SAMHD1), a deoxyribonucleoside triphosphate triphosphohydrolase that decreases dNTP pools, is frequently mutated in colon cancers, that these mutations negatively affect SAMHD1 activity, and that several SAMHD1 mutations are found in tumors with defective mismatch repair. We show that minor changes in dNTP pools in combination with inactivated mismatch repair dramatically increase mutation rates. Determination of dNTP pools in mouse embryos revealed that inactivation of one SAMHD1 allele is sufficient to elevate dNTP pools. These observations suggest that heterozygous cancer-associated SAMHD1 mutations increase mutation rates in cancer cells.
Keywords: DNA replication fidelity; colon cancer; dNTP pools.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



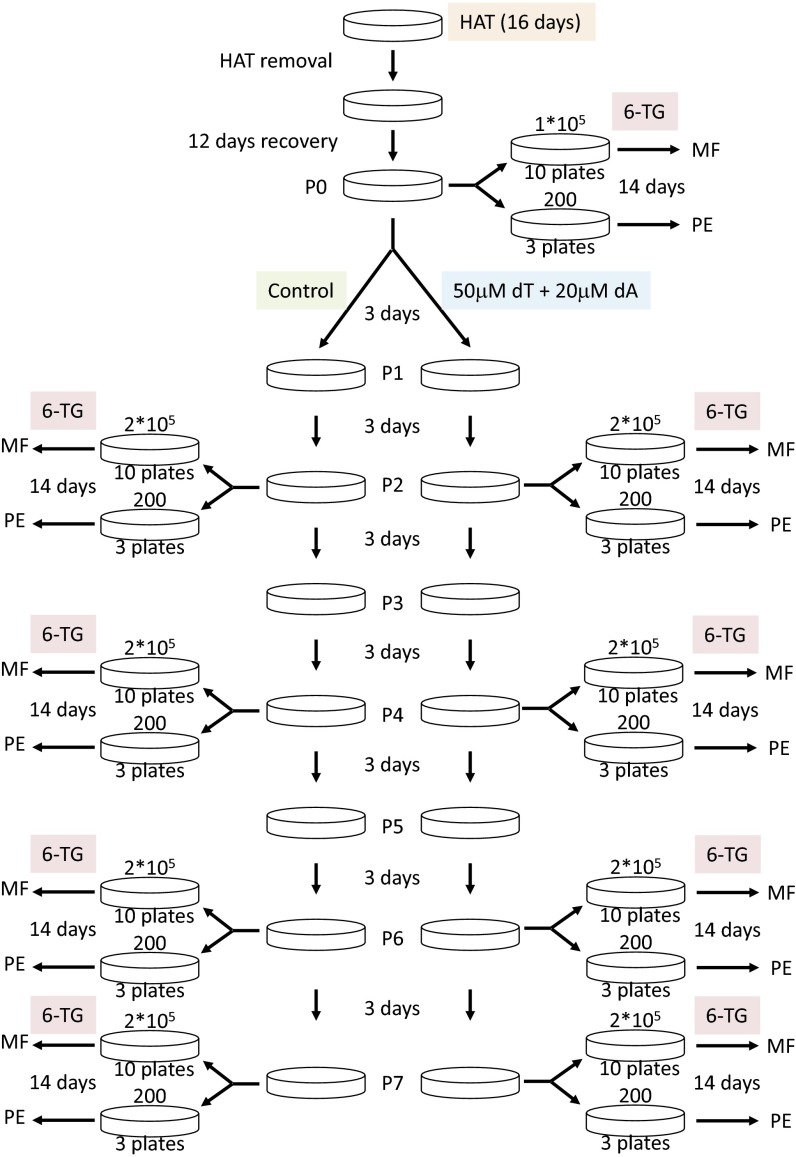
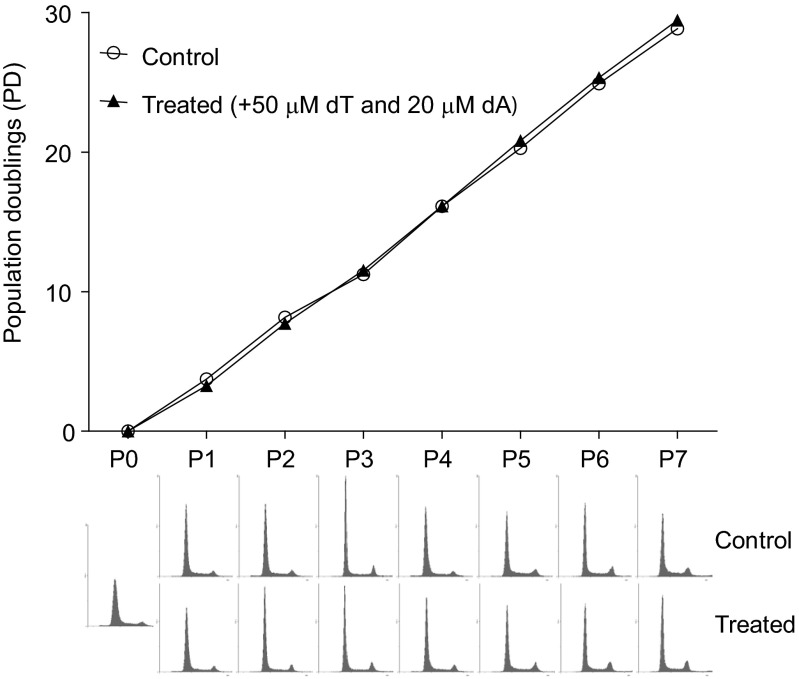
References
-
- Loeb LA, Springgate CF, Battula N. Errors in DNA replication as a basis of malignant changes. Cancer Res. 1974;34(9):2311–2321. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
