

Inhibition of translation initiation complex formation by GE81112 unravels a 16S rRNA structural switch involved in P-site decoding
- PMID: 27071098
- PMCID: PMC4843482
- DOI: 10.1073/pnas.1521156113
Inhibition of translation initiation complex formation by GE81112 unravels a 16S rRNA structural switch involved in P-site decoding
Abstract
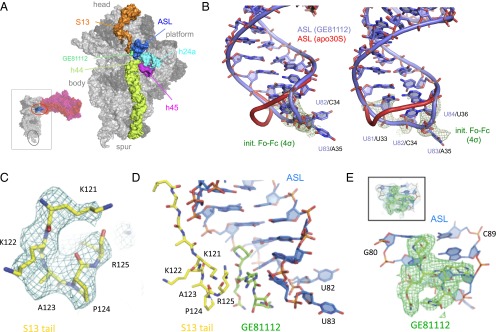
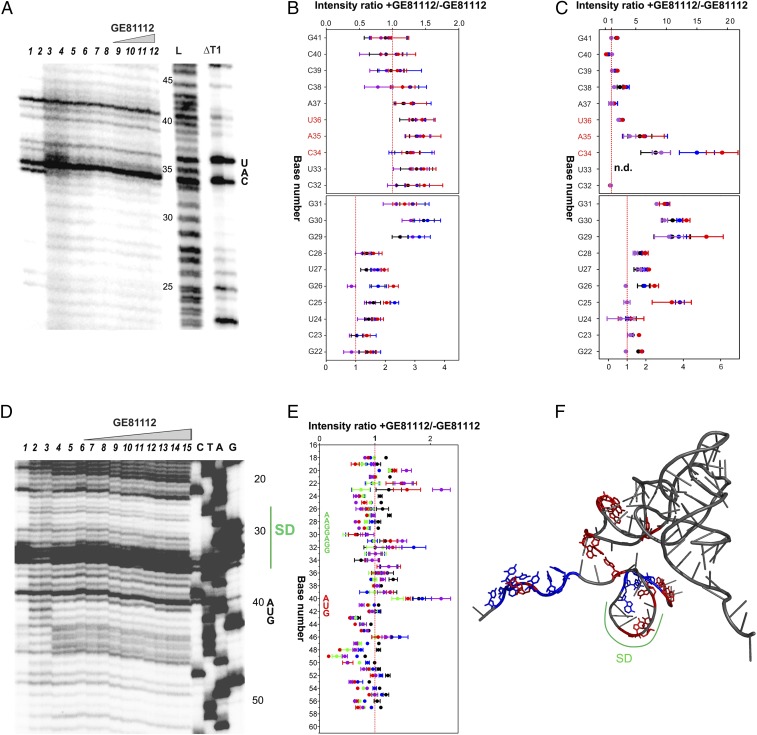
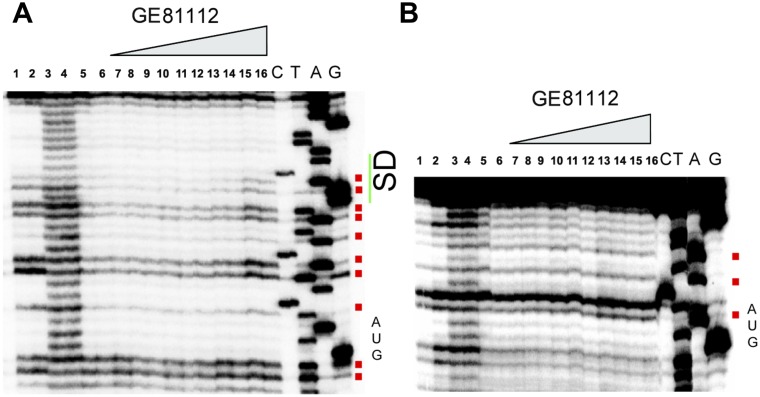
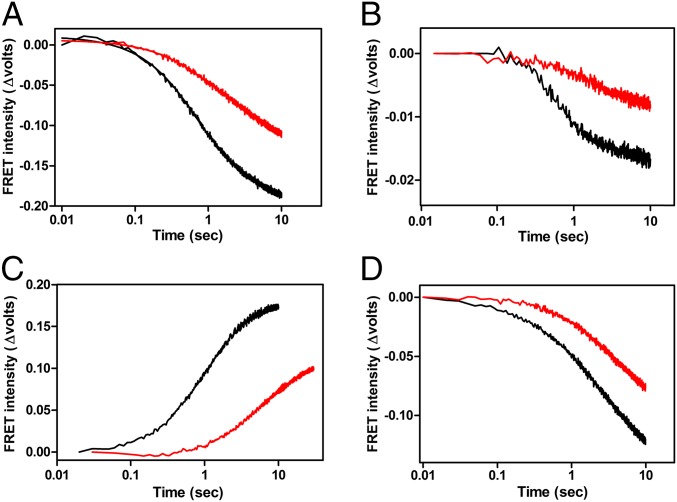
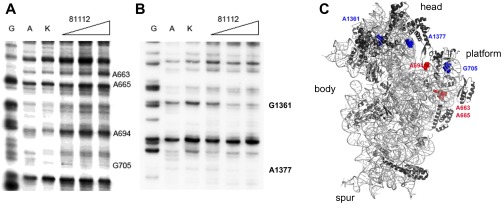
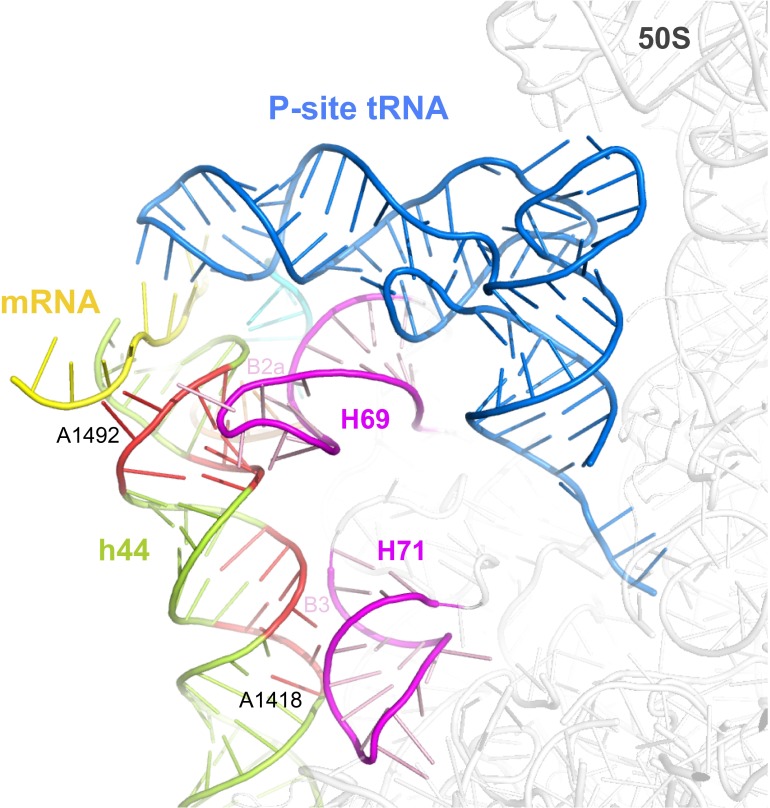
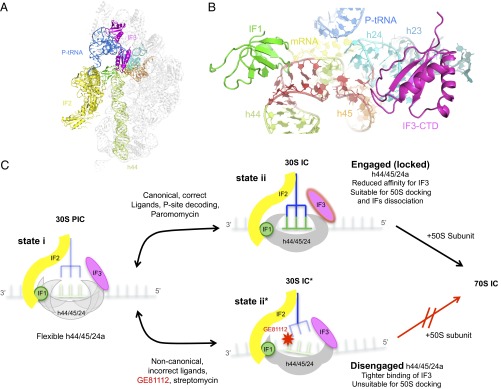
In prokaryotic systems, the initiation phase of protein synthesis is governed by the presence of initiation factors that guide the transition of the small ribosomal subunit (30S) from an unlocked preinitiation complex (30S preIC) to a locked initiation complex (30SIC) upon the formation of a correct codon-anticodon interaction in the peptidyl (P) site. Biochemical and structural characterization of GE81112, a translational inhibitor specific for the initiation phase, indicates that the main mechanism of action of this antibiotic is to prevent P-site decoding by stabilizing the anticodon stem loop of the initiator tRNA in a distorted conformation. This distortion stalls initiation in the unlocked 30S preIC state characterized by tighter IF3 binding and a reduced association rate for the 50S subunit. At the structural level we observe that in the presence of GE81112 the h44/h45/h24a interface, which is part of the IF3 binding site and forms ribosomal intersubunit bridges, preferentially adopts a disengaged conformation. Accordingly, the findings reveal that the dynamic equilibrium between the disengaged and engaged conformations of the h44/h45/h24a interface regulates the progression of protein synthesis, acting as a molecular switch that senses and couples the 30S P-site decoding step of translation initiation to the transition from an unlocked preIC to a locked 30SIC state.
Keywords: GE81112; X-ray crystallography; antibiotics; protein synthesis; ribosome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- Gale EF. The Molecular basis of Antibiotic Action. Wiley; London: 1981.
-
- Brandi L, Fabbretti A, Pon CL, Dahlberg AE, Gualerzi CO. Initiation of protein synthesis: A target for antimicrobials. Expert Opin Ther Targets. 2008;12(5):519–534. - PubMed
-
- Wilson DN. The A-Z of bacterial translation inhibitors. Crit Rev Biochem Mol Biol. 2009;44(6):393–433. - PubMed
-
- Fabbretti A, Gualerzi CO, Brandi L. How to cope with the quest for new antibiotics. FEBS Lett. 2011;585(11):1673–1681. - PubMed
-
- Brandi L, et al. The translation initiation functions of IF2: Targets for thiostrepton inhibition. J Mol Biol. 2004;335(4):881–894. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
