

The innate immune properties of airway mucosal surfaces are regulated by dynamic interactions between mucins and interacting proteins: the mucin interactome
- PMID: 27072609
- PMCID: PMC5063652
- DOI: 10.1038/mi.2016.27
The innate immune properties of airway mucosal surfaces are regulated by dynamic interactions between mucins and interacting proteins: the mucin interactome
Abstract
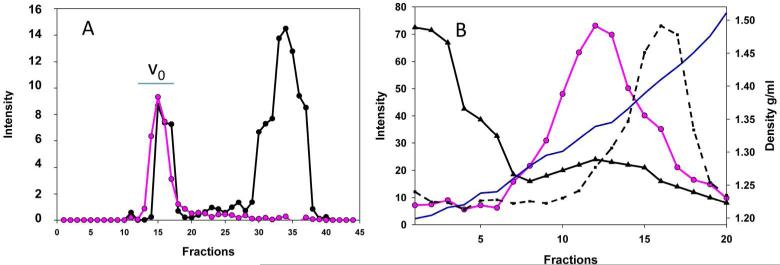
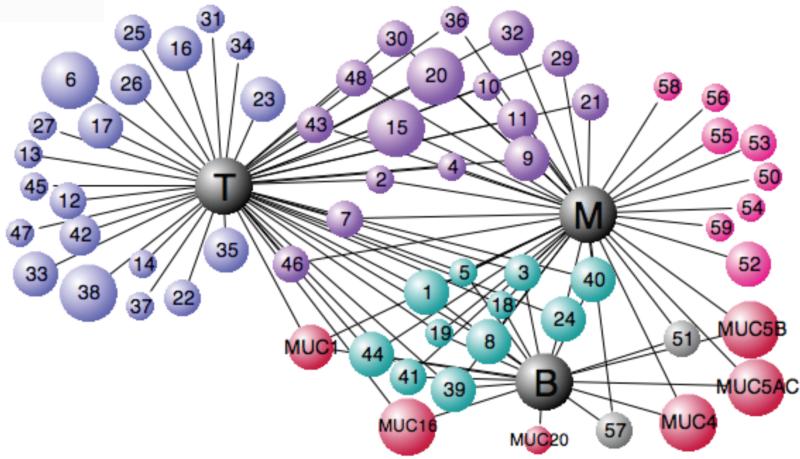
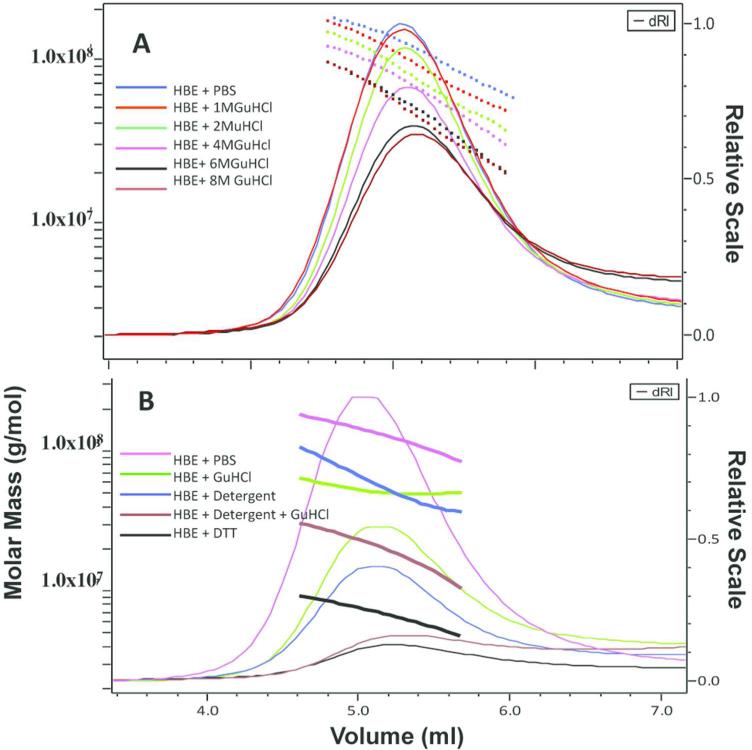
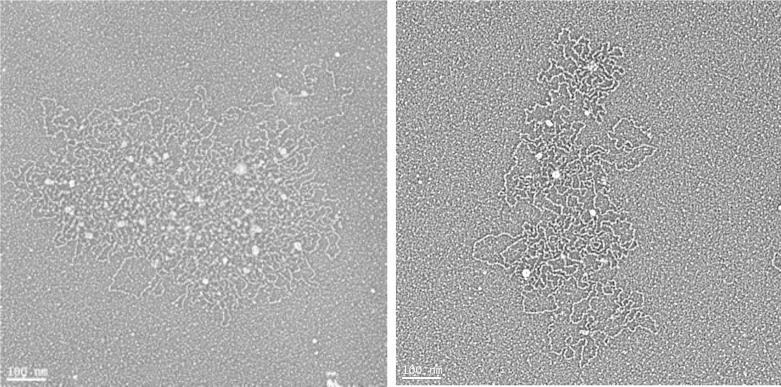
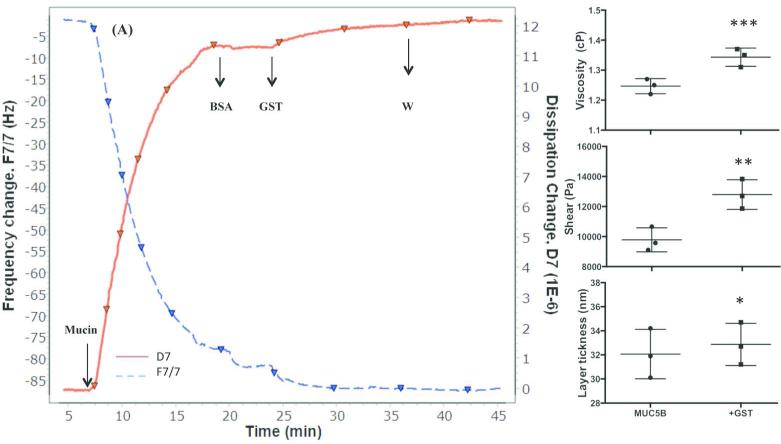
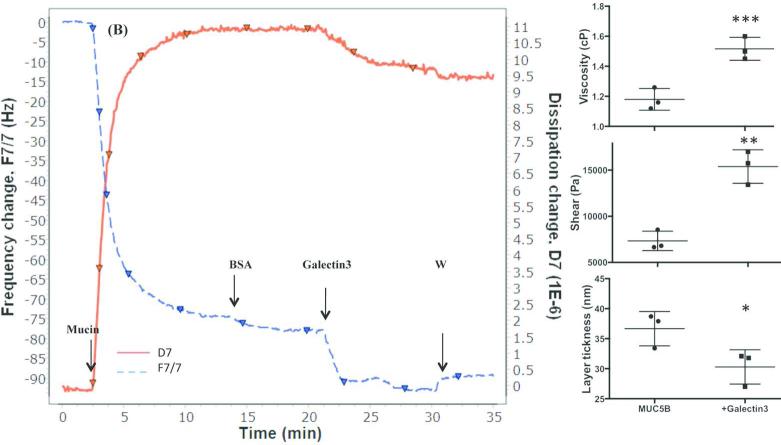
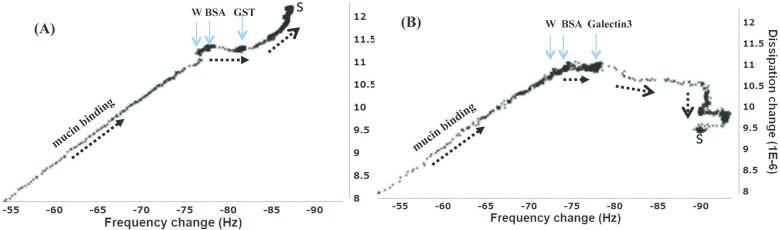
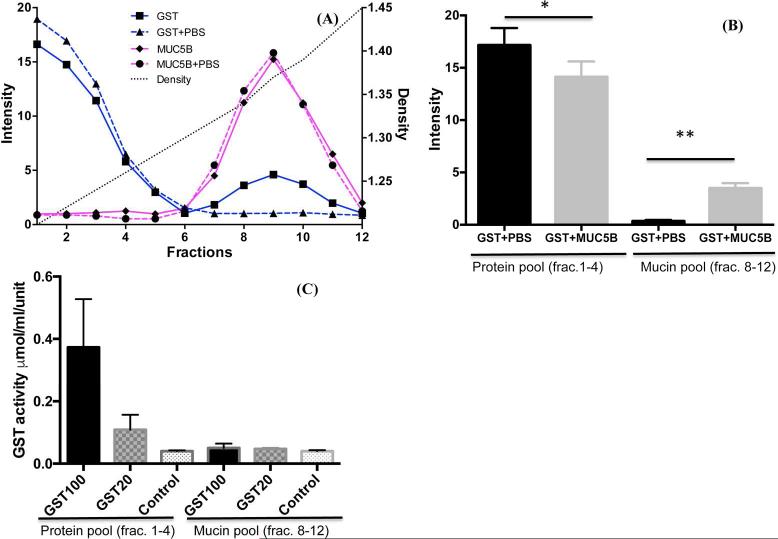
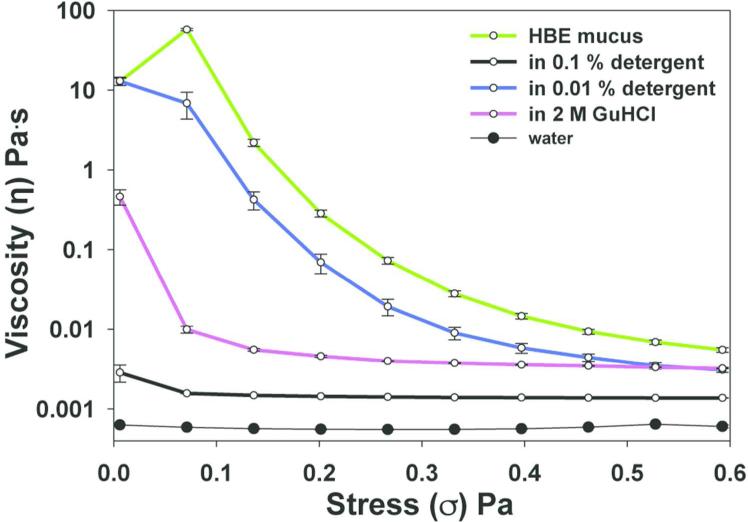
Chronic lung diseases such as cystic fibrosis, chronic bronchitis, and asthma are characterized by hypersecretion and poor clearance of mucus, which are associated with poor prognosis and mortality. Little is known about the relationship between the biophysical properties of mucus and its molecular composition. The mucins MUC5B and MUC5AC are traditionally believed to generate the characteristic biophysical properties of airway mucus. However, the contribution of hundreds of globular proteins to the biophysical properties of mucus is not clear. Approximately one-third of the total mucus proteome comprises distinct, multi-protein complexes centered around airway mucins. These complexes constitute a discrete entity we call the "mucin interactome". The data suggest that while the majority of these proteins interact with mucins via electrostatic and weak interactions, some interact through very strong hydrophobic and/or covalent interactions. Using reagents that interfere with protein-protein interactions, the complexes can be disassembled, and mucus rheology can be dramatically altered. Using MUC5B-glutathione S-transferase (GST) and MUC5B-galectin-3 as a representative of these interactions, we provide evidence that individual mucin protein interactions can alter the biophysical properties of mucus and modulate the biological function of the protein. We propose that the key mechano- and bio-active functions of mucus depend on the dynamic interactions between mucins and globular proteins. These observations challenge the paradigm that mucins are the only molecules that confer biophysical properties of mucus. These observations may ultimately lead to a greater understanding of the system and guide the development of strategies for more effective interventions using better therapeutic agents.
Figures

References
-
- Sheehan JK, Kesimer M, Pickles R. Innate immunity and mucus structure and function. Novartis Foundation symposium. 2006;279:155–166. discussion 167-159, 216-159. - PubMed
-
- Rubin BK. Physiology of airway mucus clearance. Respiratory care. 2002;47(7):761–768. - PubMed
-
- Rogers DF. Mucus hypersecretion in chronic obstructive pulmonary disease. Novartis Foundation symposium. 2001;234:65–77. discussion 77-83. - PubMed
-
- Rubin BK. Mucus structure and properties in cystic fibrosis. Paediatric respiratory reviews. 2007;8(1):4–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
