

Instant Integrated Ultradeep Quantitative-structural Membrane Proteomics Discovered Post-translational Modification Signatures for Human Cys-loop Receptor Subunit Bias
- PMID: 27073180
- PMCID: PMC5141279
- DOI: 10.1074/mcp.M114.047514
Instant Integrated Ultradeep Quantitative-structural Membrane Proteomics Discovered Post-translational Modification Signatures for Human Cys-loop Receptor Subunit Bias
Abstract
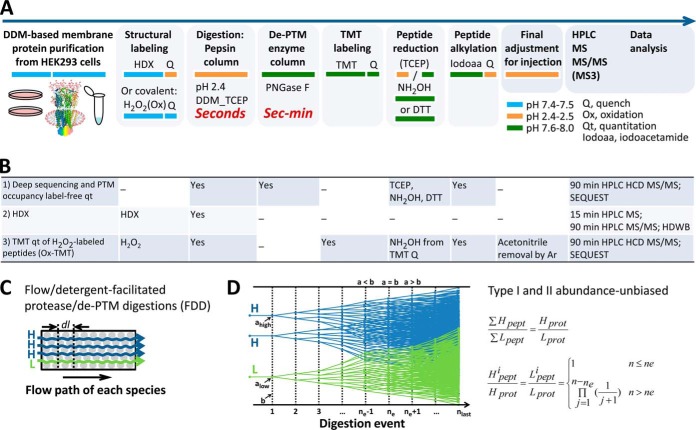
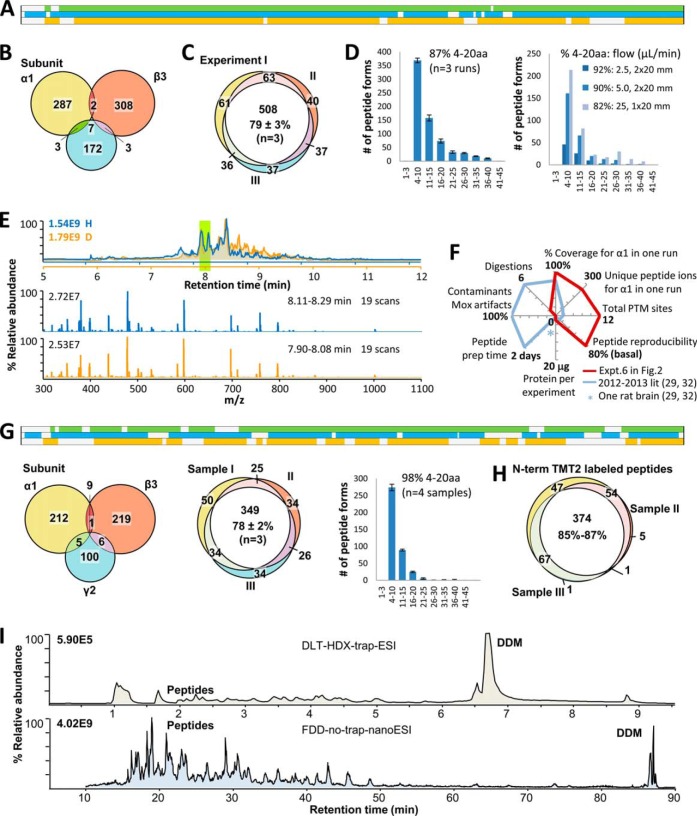
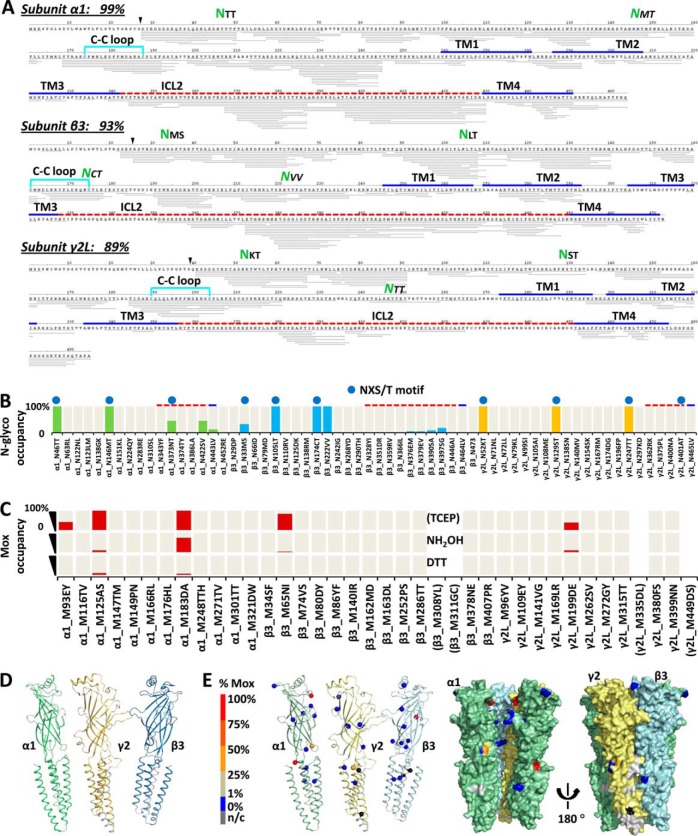
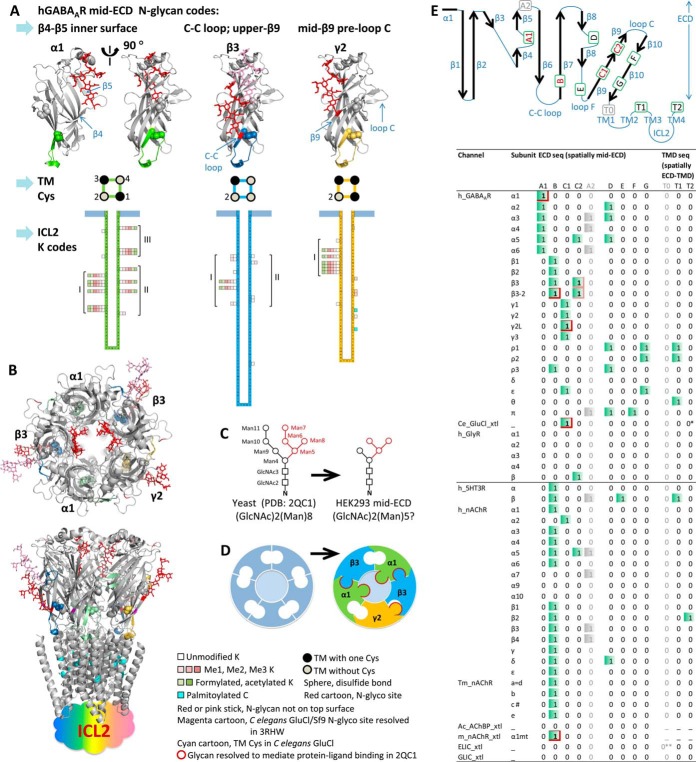
Neurotransmitter ligand-gated ion channels (LGICs) are widespread and pivotal in brain functions. Unveiling their structure-function mechanisms is crucial to drive drug discovery, and demands robust proteomic quantitation of expression, post-translational modifications (PTMs) and dynamic structures. Yet unbiased digestion of these modified transmembrane proteins-at high efficiency and peptide reproducibility-poses the obstacle. Targeting both enzyme-substrate contacts and PTMs for peptide formation and detection, we devised flow-and-detergent-facilitated protease and de-PTM digestions for deep sequencing (FDD) method that combined omni-compatible detergent, tandem immobilized protease/PNGase columns, and Cys-selective reduction/alkylation, to achieve streamlined ultradeep peptide preparation within minutes not days, at high peptide reproducibility and low abundance-bias. FDD transformed enzyme-protein contacts into equal catalytic travel paths through enzyme-excessive columns regardless of protein abundance, removed products instantly preventing inhibition, tackled intricate structures via sequential multiple micro-digestions along the flow, and precisely controlled peptide formation by flow rate. Peptide-stage reactions reduced steric bias; low contamination deepened MS/MS scan; distinguishing disulfide from M oxidation and avoiding gain/loss artifacts unmasked protein-endogenous oxidation states. Using a recent interactome of 285-kDa human GABA type A receptor, this pilot study validated FDD platform's applicability to deep sequencing (up to 99% coverage), H/D-exchange and TMT-based structural mapping. FDD discovered novel subunit-specific PTM signatures, including unusual nontop-surface N-glycosylations, that may drive subunit biases in human Cys-loop LGIC assembly and pharmacology, by redefining subunit/ligand interfaces and connecting function domains.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

References
-
- Pieper U., Schlessinger A., Kloppmann E., Chang G. A., Chou J. J., Dumont M. E., Fox B. G., Fromme P., Hendrickson W. A., Malkowski M. G., Rees D. C., Stokes D. L., Stowell M. H., Wiener M. C., Rost B., Stroud R. M., Stevens R. C., and Sali A. (2013) Coordinating the impact of structural genomics on the human alpha-helical transmembrane proteome. Nat. Struct. Mol. Biol. 20, 135–138 - PMC - PubMed
-
- Marini C., Harkin L. A., Wallace R. H., Mulley J. C., Scheffer I. E., and Berkovic S. F. (2003) Childhood absence epilepsy and febrile seizures: a family with a GABA(A) receptor mutation. Brain 126, 230–240 - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
