

The Rewiring of Ubiquitination Targets in a Pathogenic Yeast Promotes Metabolic Flexibility, Host Colonization and Virulence
- PMID: 27073846
- PMCID: PMC4830568
- DOI: 10.1371/journal.ppat.1005566
The Rewiring of Ubiquitination Targets in a Pathogenic Yeast Promotes Metabolic Flexibility, Host Colonization and Virulence
Abstract
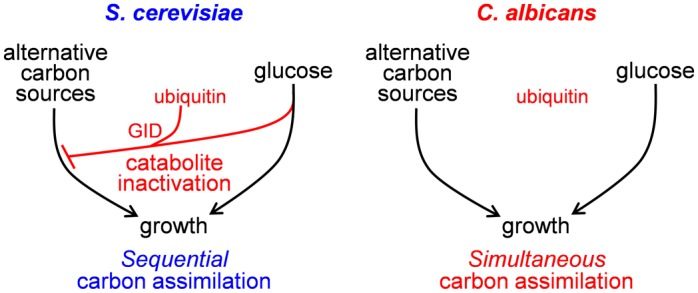
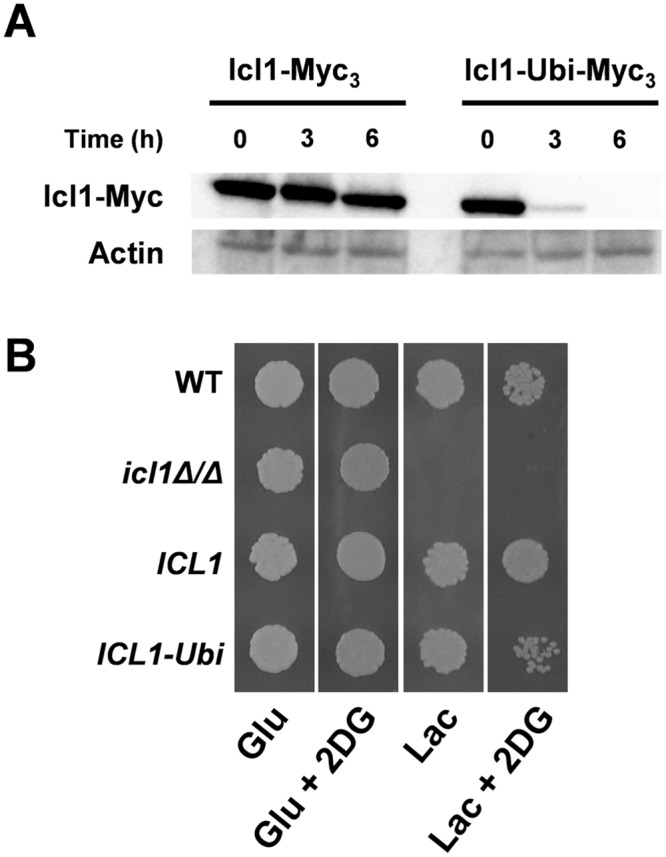
Efficient carbon assimilation is critical for microbial growth and pathogenesis. The environmental yeast Saccharomyces cerevisiae is "Crabtree positive", displaying a rapid metabolic switch from the assimilation of alternative carbon sources to sugars. Following exposure to sugars, this switch is mediated by the transcriptional repression of genes (carbon catabolite repression) and the turnover (catabolite inactivation) of enzymes involved in the assimilation of alternative carbon sources. The pathogenic yeast Candida albicans is Crabtree negative. It has retained carbon catabolite repression mechanisms, but has undergone posttranscriptional rewiring such that gluconeogenic and glyoxylate cycle enzymes are not subject to ubiquitin-mediated catabolite inactivation. Consequently, when glucose becomes available, C. albicans can continue to assimilate alternative carbon sources alongside the glucose. We show that this metabolic flexibility promotes host colonization and virulence. The glyoxylate cycle enzyme isocitrate lyase (CaIcl1) was rendered sensitive to ubiquitin-mediated catabolite inactivation in C. albicans by addition of a ubiquitination site. This mutation, which inhibits lactate assimilation in the presence of glucose, reduces the ability of C. albicans cells to withstand macrophage killing, colonize the gastrointestinal tract and cause systemic infections in mice. Interestingly, most S. cerevisiae clinical isolates we examined (67%) have acquired the ability to assimilate lactate in the presence of glucose (i.e. they have become Crabtree negative). These S. cerevisiae strains are more resistant to macrophage killing than Crabtree positive clinical isolates. Moreover, Crabtree negative S. cerevisiae mutants that lack Gid8, a key component of the Glucose-Induced Degradation complex, are more resistant to macrophage killing and display increased virulence in immunocompromised mice. Thus, while Crabtree positivity might impart a fitness advantage for yeasts in environmental niches, the more flexible carbon assimilation strategies offered by Crabtree negativity enhance the ability of yeasts to colonize and infect the mammalian host.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures








References
-
- Johnston M. Feasting, fasting and fermenting. Glucose sensing in yeast and other cells. Trends Genet. 1999;15(1):29–33. . - PubMed
-
- Inada T, Kimata K, Aiba H. Mechanism responsible for glucose-lactose diauxie in Escherichia coli: challenge to the cAMP model. Genes Cells. 1996;1(3):293–301. . - PubMed
-
- Lohr D, Venkov P, Zlatanova J. Transcriptional regulation in the yeast GAL gene family: a complex genetic network. FASEB J. 1995;9(9):777–87. . - PubMed
-
- Niimi M, Kamiyama A, Tokunaga M. Respiration of medically important Candida species and Saccharomyces cerevisiae in relation to glucose effect. J Med Vet Mycol. 1988;26(3):195–8. . - PubMed
Publication types
MeSH terms
Grants and funding
- 097377/WT_/Wellcome Trust/United Kingdom
- BB/K017365/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F00513X/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- NC/K000306/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
