

Translocator Protein 18 kDa (TSPO): An Old Protein with New Functions?
- PMID: 27074410
- PMCID: PMC5125027
- DOI: 10.1021/acs.biochem.6b00142
Translocator Protein 18 kDa (TSPO): An Old Protein with New Functions?
Abstract
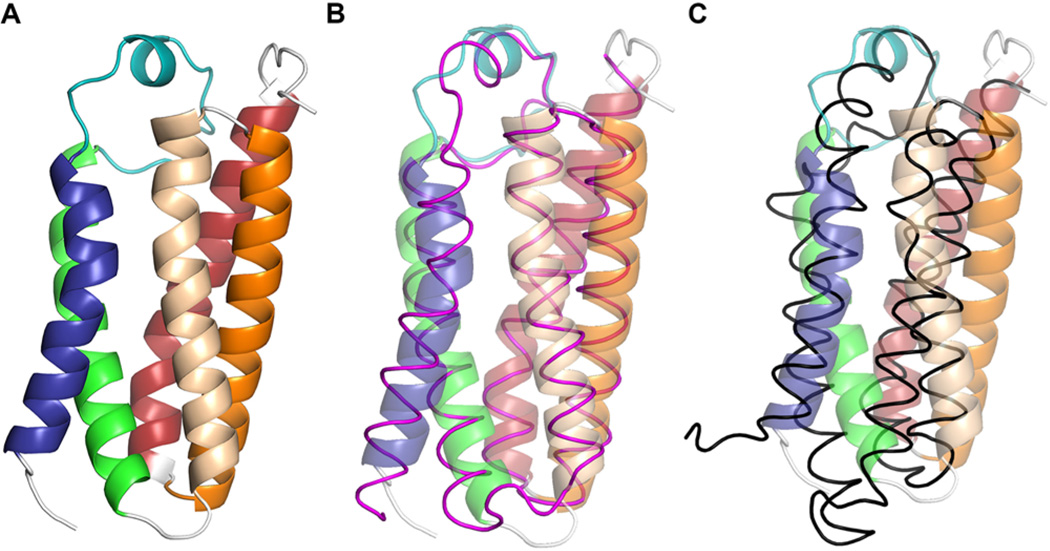
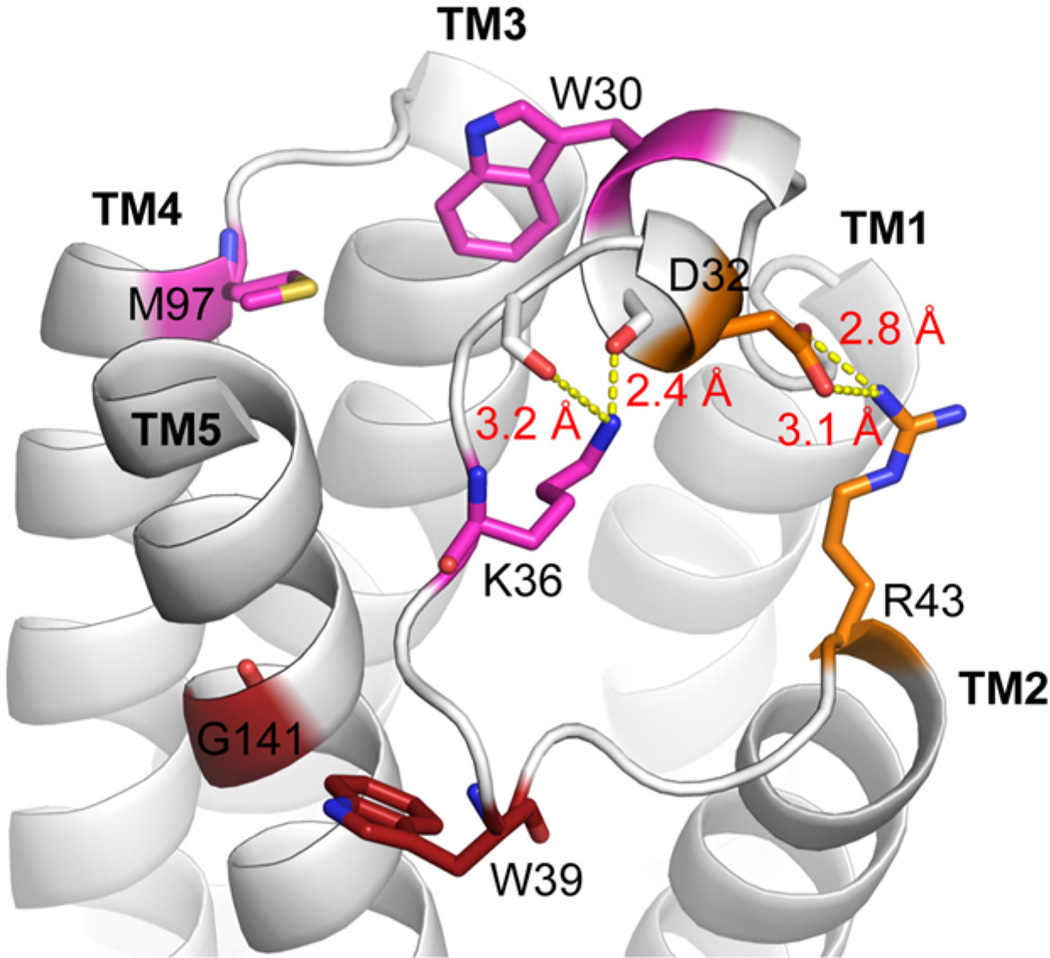
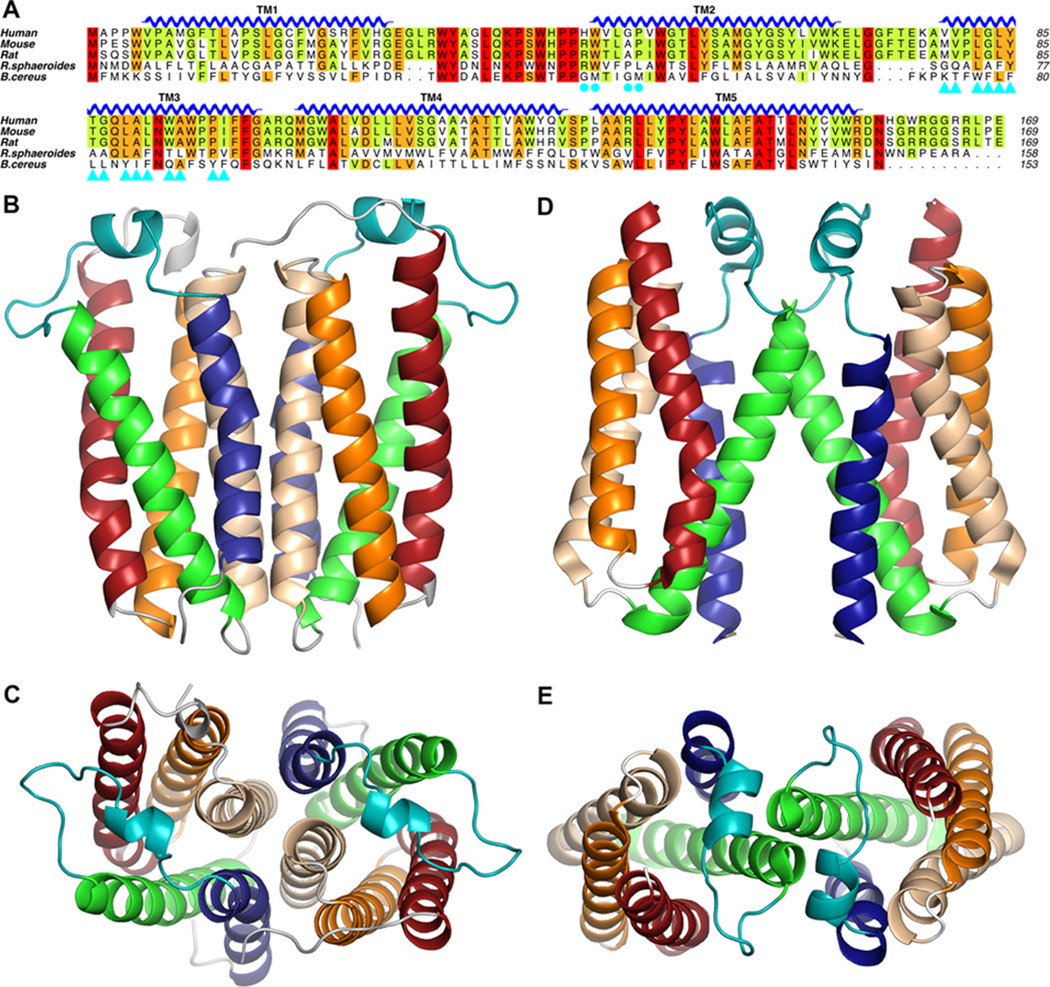
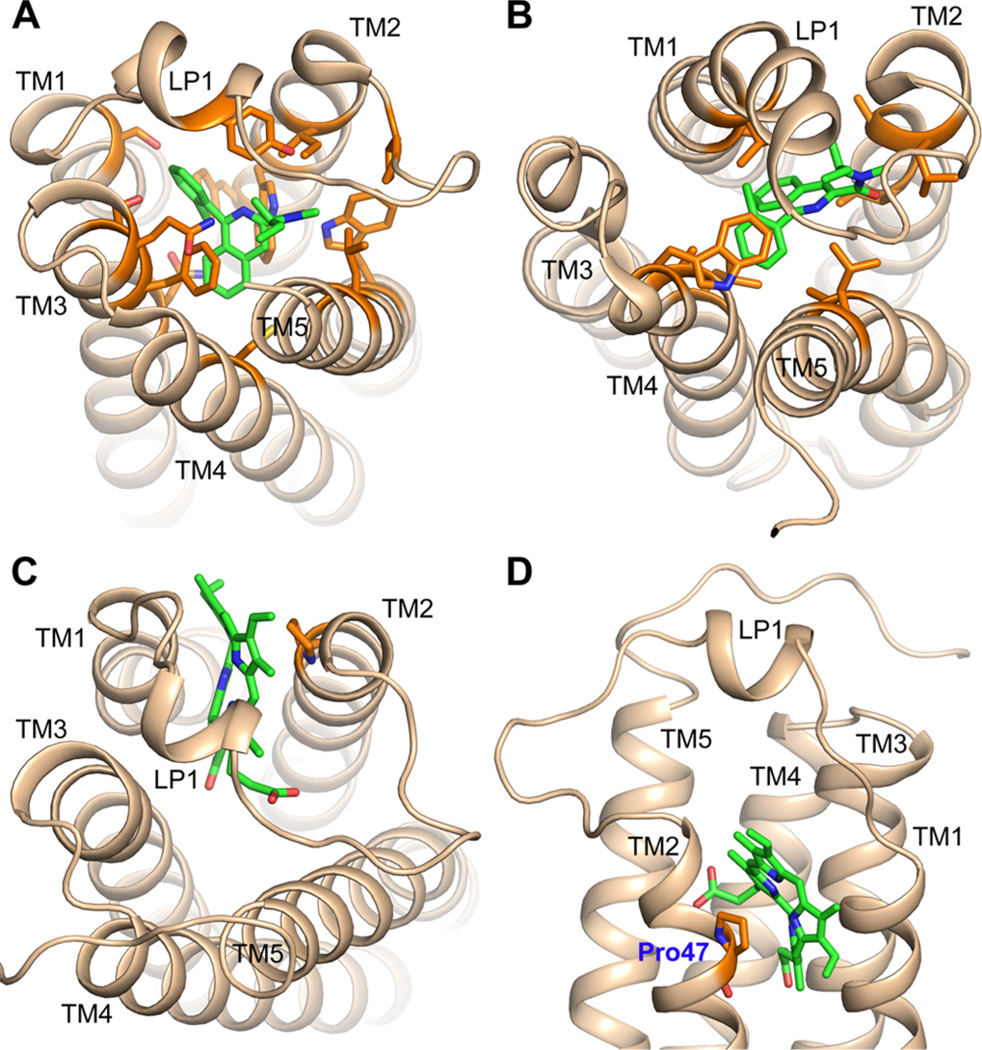
Translocator protein 18 kDa (TSPO) was previously known as the peripheral benzodiazepine receptor (PBR) in eukaryotes, where it is mainly localized to the mitochondrial outer membrane. Considerable evidence indicates that it plays regulatory roles in steroidogenesis and apoptosis and is involved in various human diseases, such as metastatic cancer, Alzheimer's and Parkinson's disease, inflammation, and anxiety disorders. Ligands of TSPO are widely used as diagnostic tools and treatment options, despite there being no clear understanding of the function of TSPO. An ortholog in the photosynthetic bacterium Rhodobacter was independently discovered as the tryptophan-rich sensory protein (TspO) and found to play a role in the response to changes in oxygen and light conditions that regulate photosynthesis and respiration. As part of this highly conserved protein family found in all three kingdoms, the rat TSPO is able to rescue the knockout phenotype in Rhodobacter, indicating functional as well as structural conservation. Recently, a major breakthrough in the field was achieved: the determination of atomic-resolution structures of TSPO from different species by several independent groups. This now allows us to reexamine the function of TSPO with a molecular perspective. In this review, we focus on recently determined structures of TSPO and their implications for potential functions of this ubiquitous multifaceted protein. We suggest that TSPO is an ancient bacterial receptor/stress sensor that has developed additional interactions, partners, and roles in its mitochondrial outer membrane environment in eukaryotes.
Figures
Similar articles
-
Structural requirements to obtain highly potent and selective 18 kDa Translocator Protein (TSPO) Ligands.Curr Top Med Chem. 2011;11(7):860-86. doi: 10.2174/156802611795165142. Curr Top Med Chem. 2011. PMID: 21291396 Review.
-
Evolving understanding of translocator protein 18 kDa (TSPO).Pharmacol Res. 2015 Sep;99:404-9. doi: 10.1016/j.phrs.2015.03.022. Epub 2015 Apr 13. Pharmacol Res. 2015. PMID: 25882248 Free PMC article. Review.
-
TSPO: An Evolutionarily Conserved Protein with Elusive Functions.Int J Mol Sci. 2018 Jun 7;19(6):1694. doi: 10.3390/ijms19061694. Int J Mol Sci. 2018. PMID: 29875327 Free PMC article. Review.
-
An update into the medicinal chemistry of translocator protein (TSPO) ligands.Eur J Med Chem. 2021 Jan 1;209:112924. doi: 10.1016/j.ejmech.2020.112924. Epub 2020 Oct 14. Eur J Med Chem. 2021. PMID: 33081988 Review.
-
The mitochondrial translocator protein (TSPO): a key multifunctional molecule in the nervous system.Biochem J. 2022 Jul 15;479(13):1455-1466. doi: 10.1042/BCJ20220050. Biochem J. 2022. PMID: 35819398 Review.
Cited by
-
Insight into the Structural Features of TSPO: Implications for Drug Development.Trends Pharmacol Sci. 2020 Feb;41(2):110-122. doi: 10.1016/j.tips.2019.11.005. Epub 2019 Dec 18. Trends Pharmacol Sci. 2020. PMID: 31864680 Free PMC article. Review.
-
Structural Prediction of the Dimeric Form of the Mammalian Translocator Membrane Protein TSPO: A Key Target for Brain Diagnostics.Int J Mol Sci. 2018 Aug 31;19(9):2588. doi: 10.3390/ijms19092588. Int J Mol Sci. 2018. PMID: 30200318 Free PMC article.
-
Inhibitory Effects of the Two Novel TSPO Ligands 2-Cl-MGV-1 and MGV-1 on LPS-induced Microglial Activation.Cells. 2019 May 22;8(5):486. doi: 10.3390/cells8050486. Cells. 2019. PMID: 31121852 Free PMC article.
-
Midazolam inhibits chondrogenesis via peripheral benzodiazepine receptor in human mesenchymal stem cells.J Cell Mol Med. 2018 May;22(5):2896-2907. doi: 10.1111/jcmm.13584. Epub 2018 Mar 7. J Cell Mol Med. 2018. PMID: 29516686 Free PMC article.
-
Genomic Evidence for Bacterial Determinants Influencing Obesity Development.Int J Environ Res Public Health. 2017 Mar 26;14(4):345. doi: 10.3390/ijerph14040345. Int J Environ Res Public Health. 2017. PMID: 28346358 Free PMC article.
References
-
- Verma A, Snyder SH. Peripheral type benzodiazepine receptors. Annu. Rev. Pharmacol. Toxicol. 1989;29:307–322. - PubMed
-
- Krueger KE, Papadopoulos V. Mitochondrial benzodiazepine receptors and the regulation of steroid biosynthesis. Annu. Rev. Pharmacol. Toxicol. 1992;32:211–237. - PubMed
-
- Wendler G, Lindemann P, Lacapère JJ, Papadopoulos V. Protoporphyrin IX binding and transport by recombinant mouse PBR. Biochem. Biophys. Res. Commun. 2003;311:847–852. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
