

Cholesterol Increases the Openness of SNARE-Mediated Flickering Fusion Pores
- PMID: 27074679
- PMCID: PMC4833774
- DOI: 10.1016/j.bpj.2016.02.019
Cholesterol Increases the Openness of SNARE-Mediated Flickering Fusion Pores
Abstract
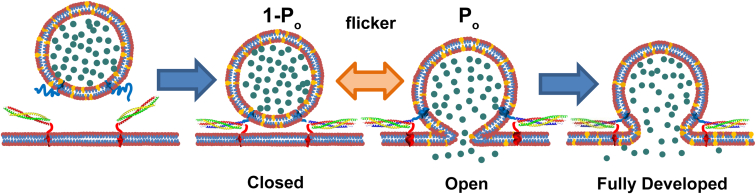
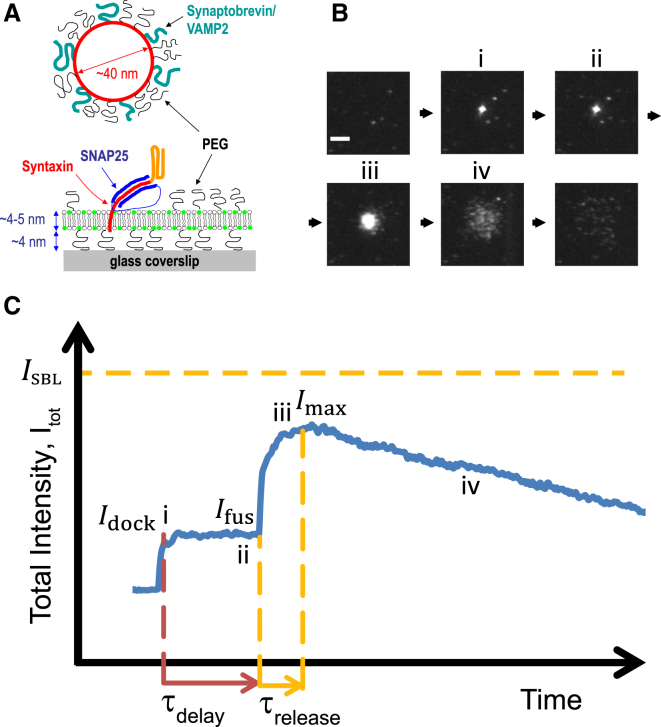
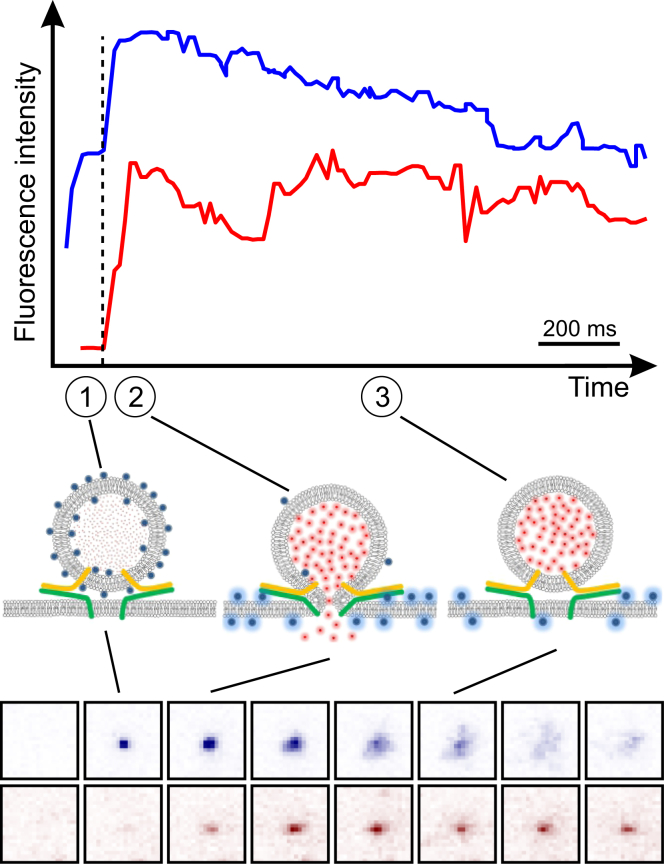
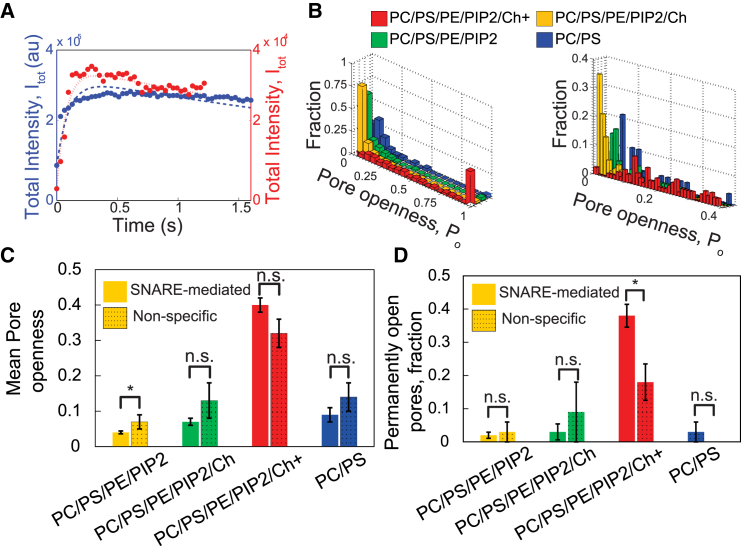
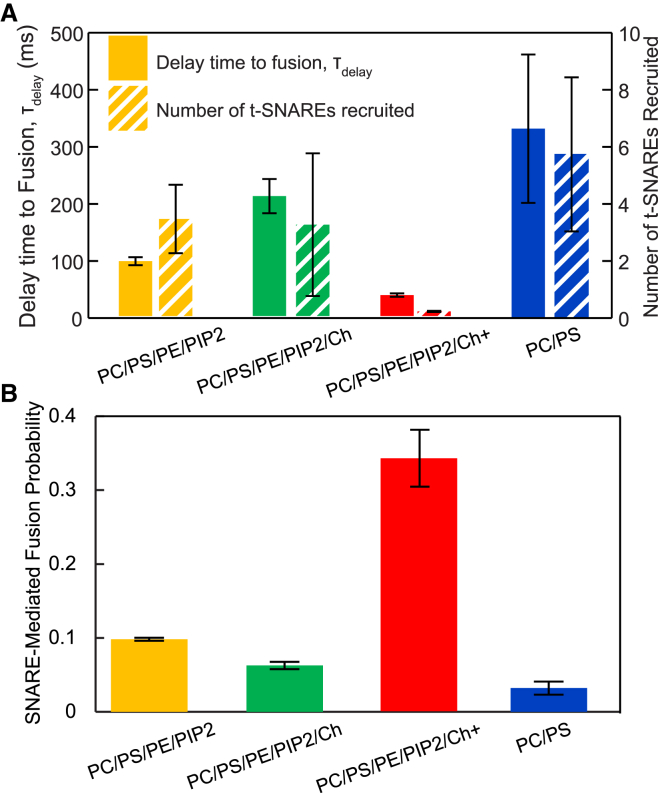
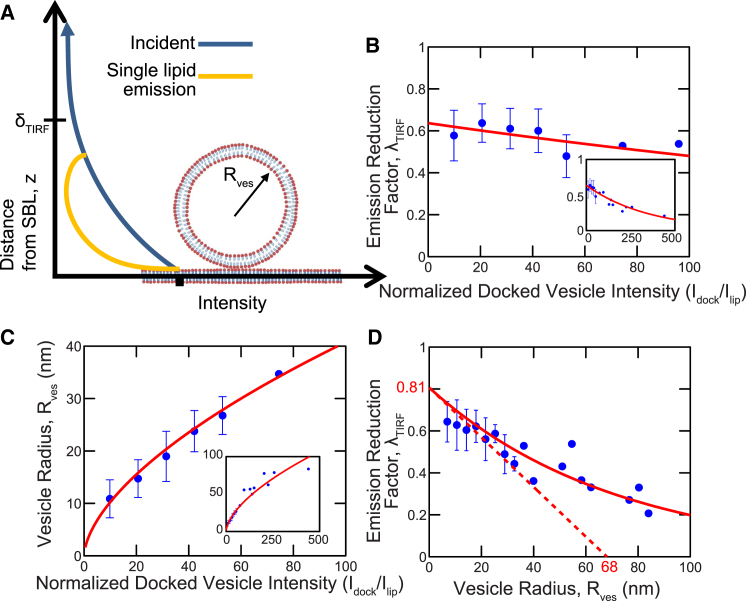
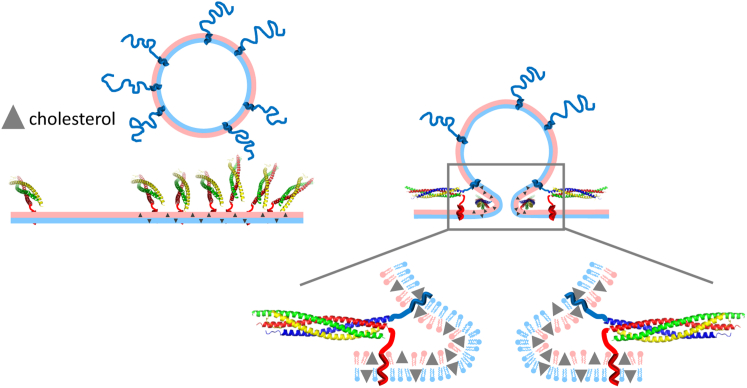
Flickering of fusion pores during exocytotic release of hormones and neurotransmitters is well documented, but without assays that use biochemically defined components and measure single-pore dynamics, the mechanisms remain poorly understood. We used total internal reflection fluorescence microscopy to quantify fusion-pore dynamics in vitro and to separate the roles of soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) proteins and lipid bilayer properties. When small unilamellar vesicles bearing neuronal v-SNAREs fused with planar bilayers reconstituted with cognate t-SNARES, lipid and soluble cargo transfer rates were severely reduced, suggesting that pores flickered. From the lipid release times we computed pore openness, the fraction of time the pore is open, which increased dramatically with cholesterol. For most lipid compositions tested, SNARE-mediated and nonspecifically nucleated pores had similar openness, suggesting that pore flickering was controlled by lipid bilayer properties. However, with physiological cholesterol levels, SNAREs substantially increased the fraction of fully open pores and fusion was so accelerated that there was insufficient time to recruit t-SNAREs to the fusion site, consistent with t-SNAREs being preclustered by cholesterol into functional docking and fusion platforms. Our results suggest that cholesterol opens pores directly by reducing the fusion-pore bending energy, and indirectly by concentrating several SNAREs into individual fusion events.
Copyright © 2016 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Lindau M., Alvarez de Toledo G. The fusion pore. Biochim. Biophys. Acta. 2003;1641:167–173. - PubMed
-
- Staal R.G.W., Mosharov E.V., Sulzer D. Dopamine neurons release transmitter via a flickering fusion pore. Nat. Neurosci. 2004;7:341–346. - PubMed
-
- Bai J., Wang C.T., Chapman E.R. Fusion pore dynamics are regulated by synaptotagmin∗t-SNARE interactions. Neuron. 2004;41:929–942. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
