

Spleen hypoplasia leads to abnormal stress hematopoiesis in mice with loss of Pbx homeoproteins in splenic mesenchyme
- PMID: 27075259
- PMCID: PMC5341595
- DOI: 10.1111/joa.12479
Spleen hypoplasia leads to abnormal stress hematopoiesis in mice with loss of Pbx homeoproteins in splenic mesenchyme
Abstract
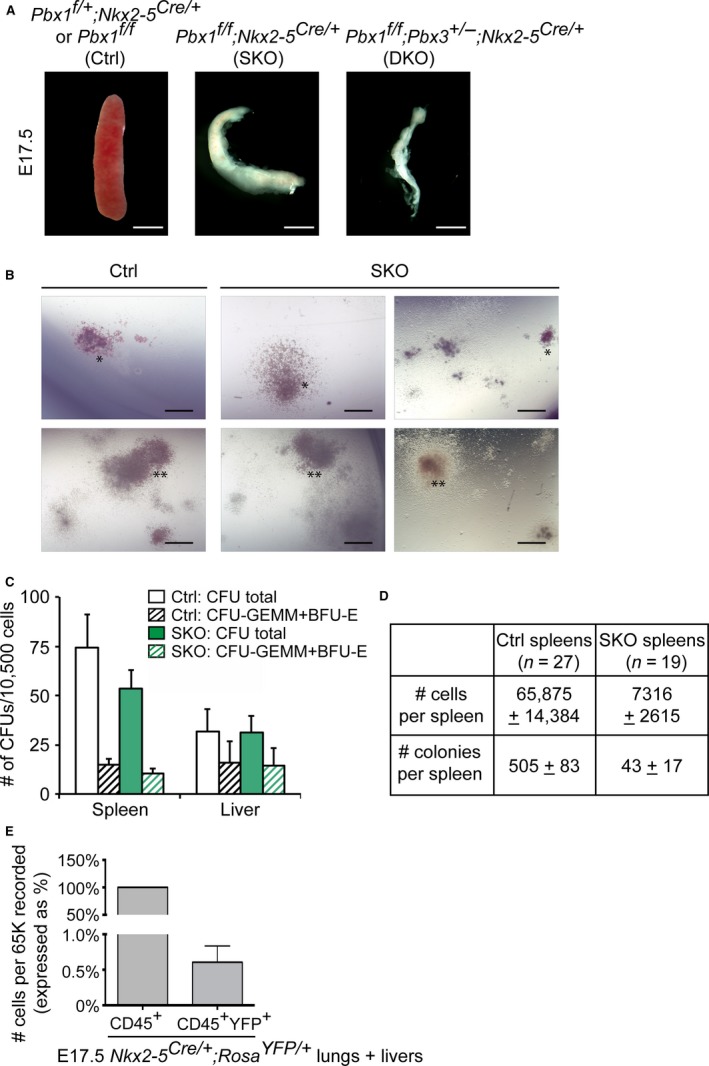
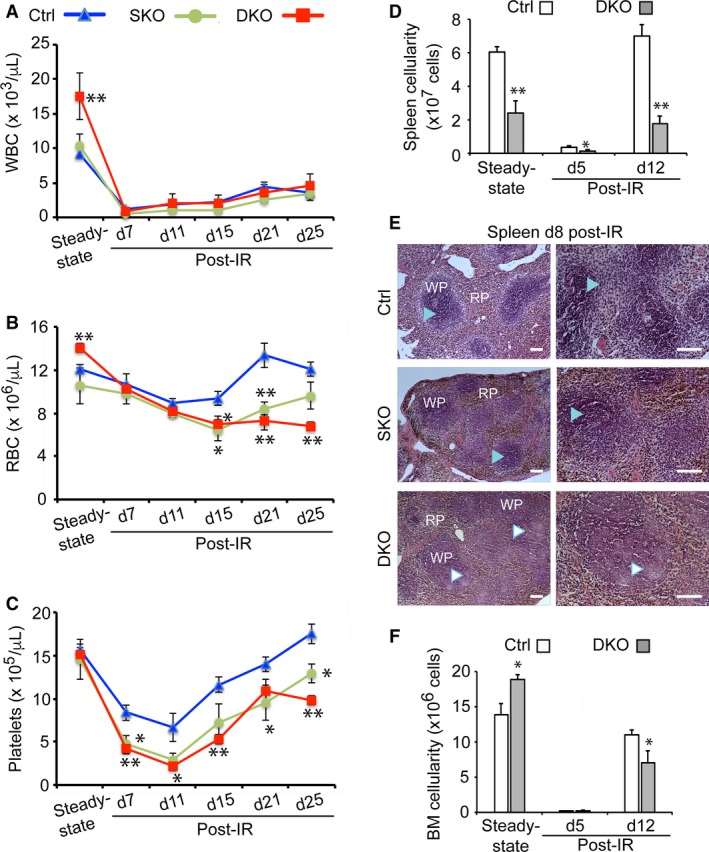
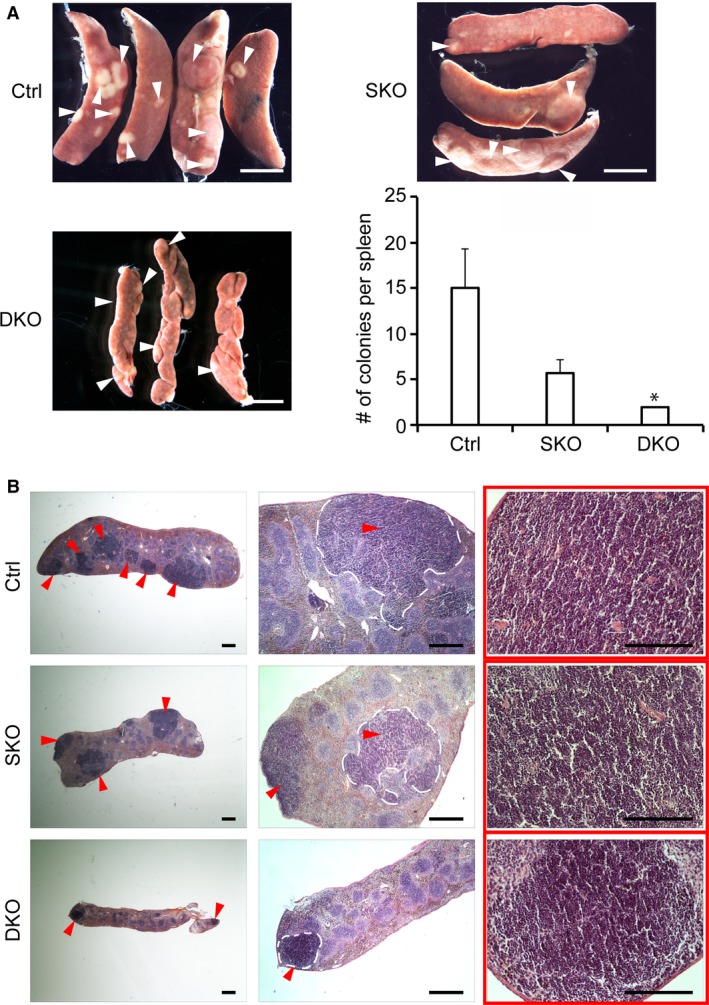
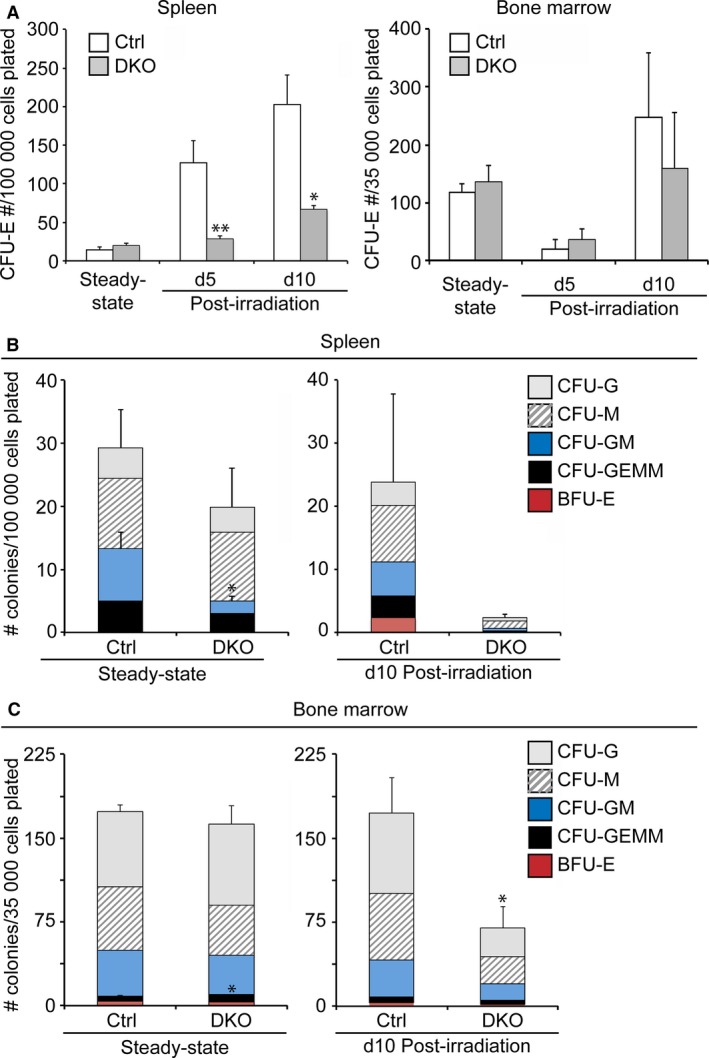
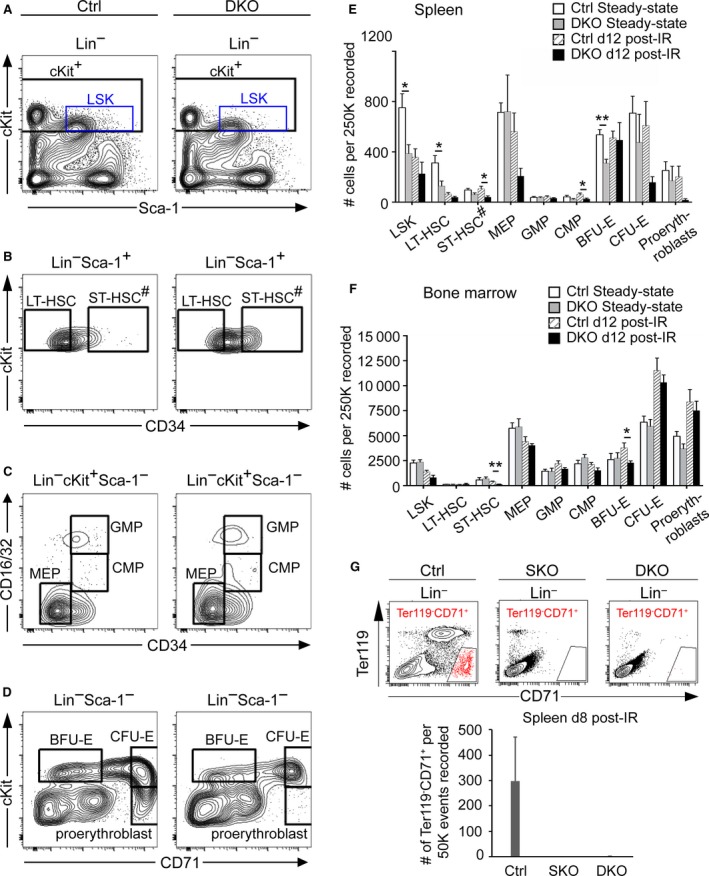
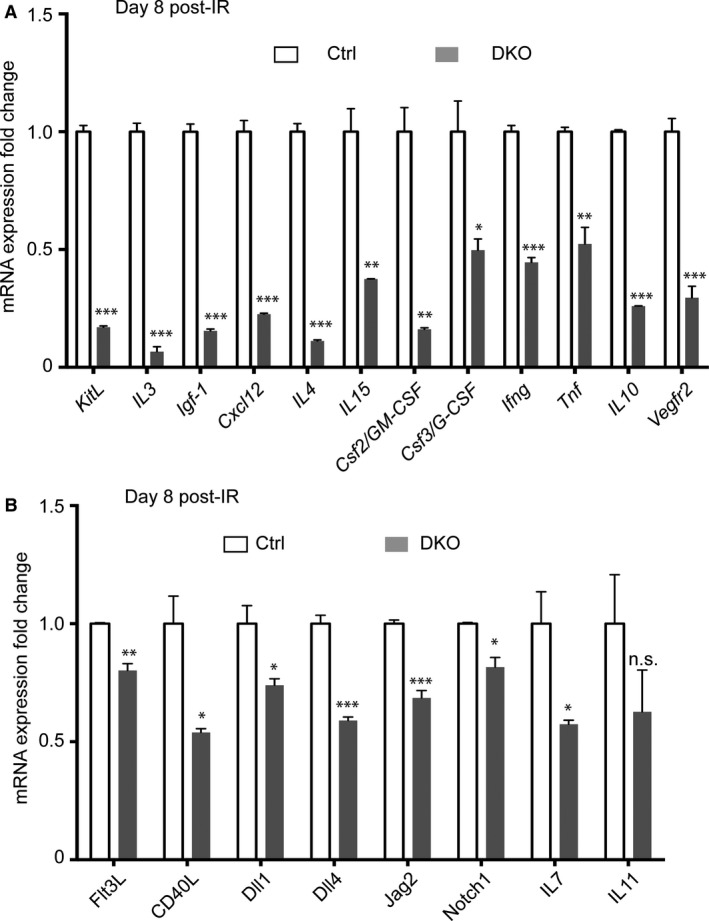
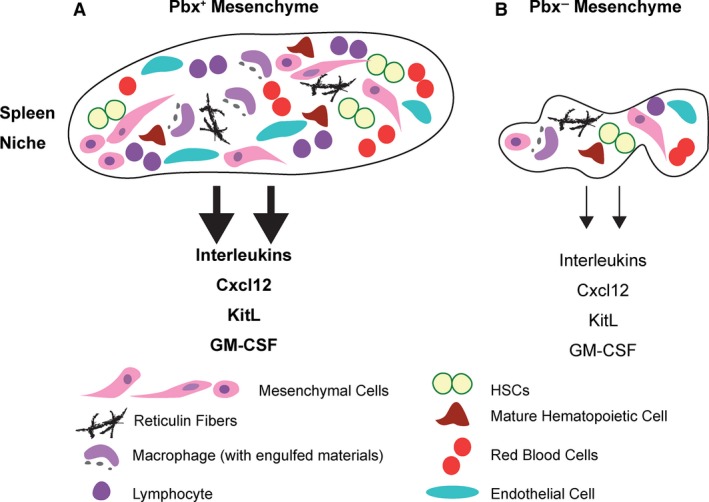
The spleen plays critical roles in immunity and also provides a permissive microenvironment for hematopoiesis. Previous studies have reported that the TALE-class homeodomain transcription factor Pbx1 is essential in hematopoietic stem and progenitor cells (HSPCs) for stem cell maintenance and progenitor expansion. However, the role of Pbx1 in the hematopoietic niche has not been investigated. Here we explored the effects that genetic perturbation of the splenic mesenchymal niche has on hematopoiesis upon loss of members of the Pbx family of homeoproteins. Splenic mesenchyme-specific inactivation of Pbx1 (SKO) on a Pbx2- or Pbx3-deficient genetic background (DKO) resulted in abnormal development of the spleen, which is dysmorphic and severely hypoplastic. This phenotype, in turn, affected the number of HSPCs in the fetal and adult spleen at steady state, as well as markedly impairing the kinetics of hematopoietic regeneration in adult mice after sub-lethal and lethal myelosuppressive irradiation. Spleens of mice with compound Pyx deficiency 8 days following sublethal irradiation displayed significant downregulation of multiple cytokine-encoding genes, including KitL/SCF, Cxcl12/SDF-1, IL-3, IL-4, GM-CSF/Csf2 IL-10, and Igf-1, compared with controls. KitL/SCF and Cxcl12/SDF-1 were recently shown to play key roles in the splenic niche in response to various haematopoietic stresses such as myeloablation, blood loss, or pregnancy. Our results demonstrate that, in addition to their intrinsic roles in HSPCs, non-cell autonomous functions of Pbx factors within the splenic niche contribute to the regulation of hematopoiesis, at least in part via the control of KitL/SCF and Cxcl12/SDF-1. Furthermore, our study establishes that abnormal spleen development and hypoplasia have deleterious effects on the efficiency of hematopoietic recovery after bone marrow injury.
Keywords: Cxcl12/SDF-1; GM-CSF; Nkx2-5+ spleen niche KitL/SCF; Pbx transcription factors; extramedullary hematopoiesis; hematopoietic stem cells; spleen hypoplasia.
© 2016 Anatomical Society.
Figures
Similar articles
-
Niche-induced extramedullary hematopoiesis in the spleen is regulated by the transcription factor Tlx1.Sci Rep. 2018 May 29;8(1):8308. doi: 10.1038/s41598-018-26693-x. Sci Rep. 2018. PMID: 29844356 Free PMC article.
-
Pbx loss in cranial neural crest, unlike in epithelium, results in cleft palate only and a broader midface.J Anat. 2018 Aug;233(2):222-242. doi: 10.1111/joa.12821. Epub 2018 May 23. J Anat. 2018. PMID: 29797482 Free PMC article.
-
A perisinusoidal niche for extramedullary haematopoiesis in the spleen.Nature. 2015 Nov 26;527(7579):466-471. doi: 10.1038/nature15530. Epub 2015 Nov 16. Nature. 2015. PMID: 26570997 Free PMC article.
-
Targeting the Spleen as an Alternative Site for Hematopoiesis.Bioessays. 2019 May;41(5):e1800234. doi: 10.1002/bies.201800234. Epub 2019 Apr 10. Bioessays. 2019. PMID: 30970171 Review.
-
Expression of FLT3 receptor and response to FLT3 ligand by leukemic cells.Leukemia. 1996 Apr;10(4):588-99. Leukemia. 1996. PMID: 8618433 Review.
Cited by
-
Computational Identification of Master Regulators Influencing Trypanotolerance in Cattle.Int J Mol Sci. 2021 Jan 8;22(2):562. doi: 10.3390/ijms22020562. Int J Mol Sci. 2021. PMID: 33429951 Free PMC article.
-
Comprehensive summary: the role of PBX1 in development and cancers.Front Cell Dev Biol. 2024 Jul 26;12:1442052. doi: 10.3389/fcell.2024.1442052. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39129784 Free PMC article. Review.
-
The regulation of PBXs and their emerging role in cancer.J Cell Mol Med. 2022 Mar;26(5):1363-1379. doi: 10.1111/jcmm.17196. Epub 2022 Jan 23. J Cell Mol Med. 2022. PMID: 35068042 Free PMC article. Review.
-
'Building a perfect body': control of vertebrate organogenesis by PBX-dependent regulatory networks.Genes Dev. 2019 Mar 1;33(5-6):258-275. doi: 10.1101/gad.318774.118. Genes Dev. 2019. PMID: 30824532 Free PMC article. Review.
-
Niche-induced extramedullary hematopoiesis in the spleen is regulated by the transcription factor Tlx1.Sci Rep. 2018 May 29;8(1):8308. doi: 10.1038/s41598-018-26693-x. Sci Rep. 2018. PMID: 29844356 Free PMC article.
References
-
- Avecilla ST, Hattori K, Heissig B, et al. (2004) Chemokine‐mediated interaction of hematopoietic progenitors with the bone marrow vascular niche is required for thrombopoiesis. Nat Med 10, 64–71. - PubMed
-
- Bertrand JY, Desanti GE, Lo‐Man R, et al. (2006) Fetal spleen stroma drives macrophage commitment. Development 133, 3619–3628. - PubMed
-
- Brendolan A, Ferretti E, Salsi V, et al. (2005) A Pbx1‐dependent genetic and transcriptional network regulates spleen ontogeny. Development 132, 3113–3126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
