

Gi-protein-coupled 5-HT1B/D receptor agonist sumatriptan induces type I hyperalgesic priming
- PMID: 27075428
- PMCID: PMC5048915
- DOI: 10.1097/j.pain.0000000000000581
Gi-protein-coupled 5-HT1B/D receptor agonist sumatriptan induces type I hyperalgesic priming
Abstract
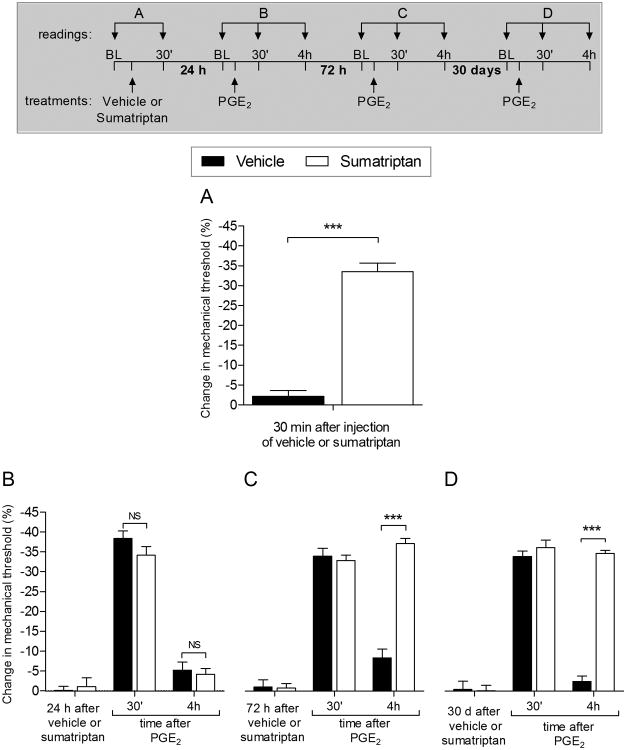
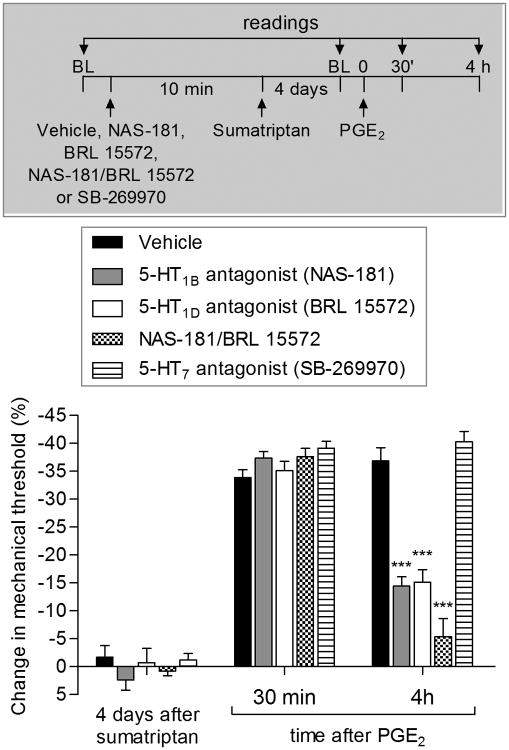
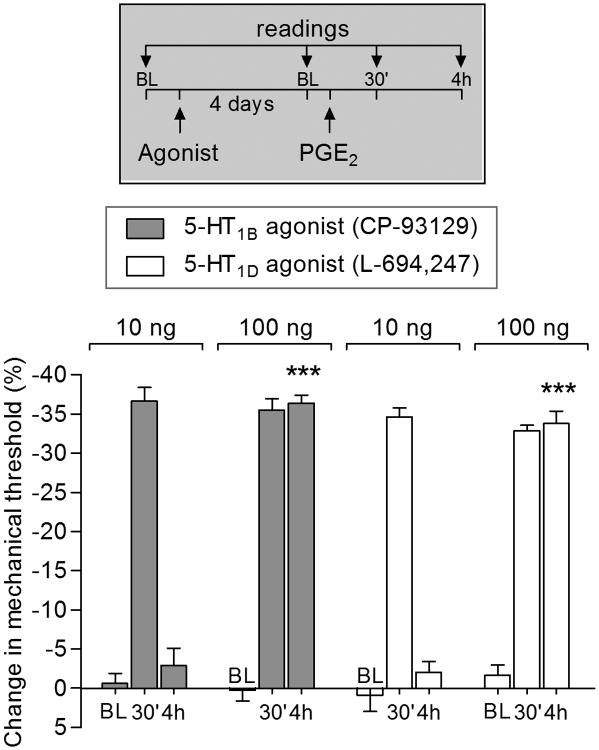
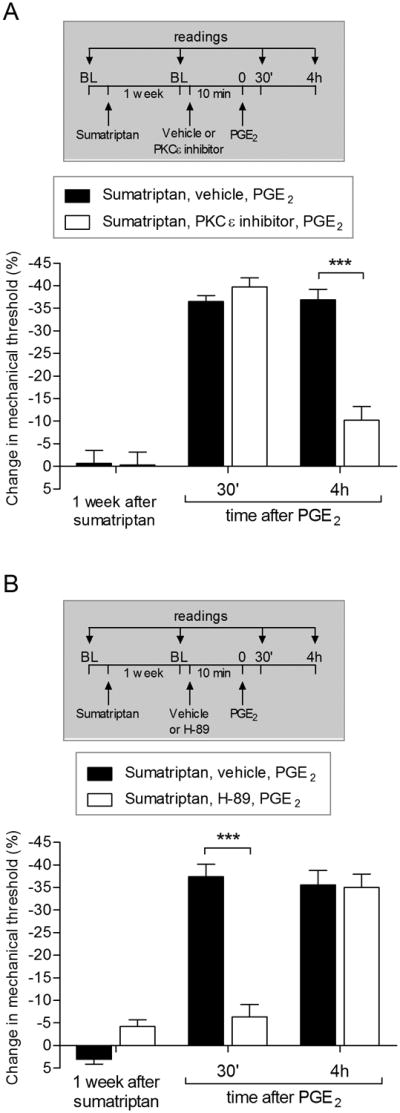
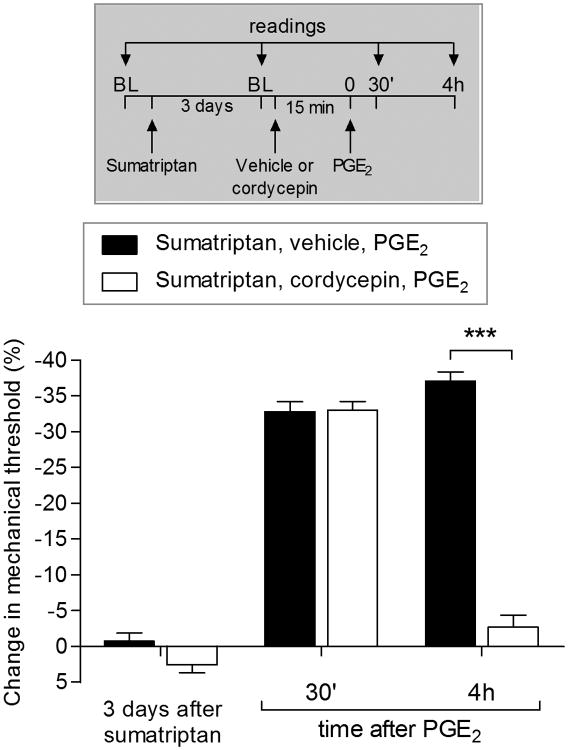
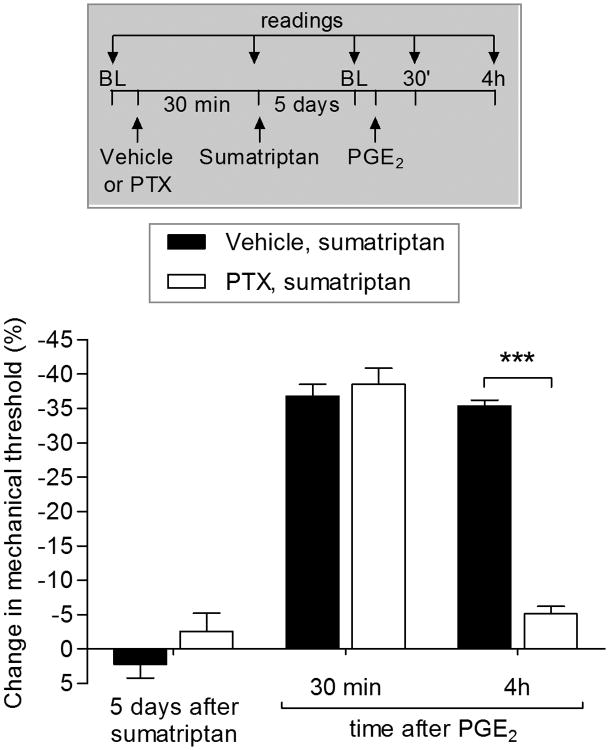
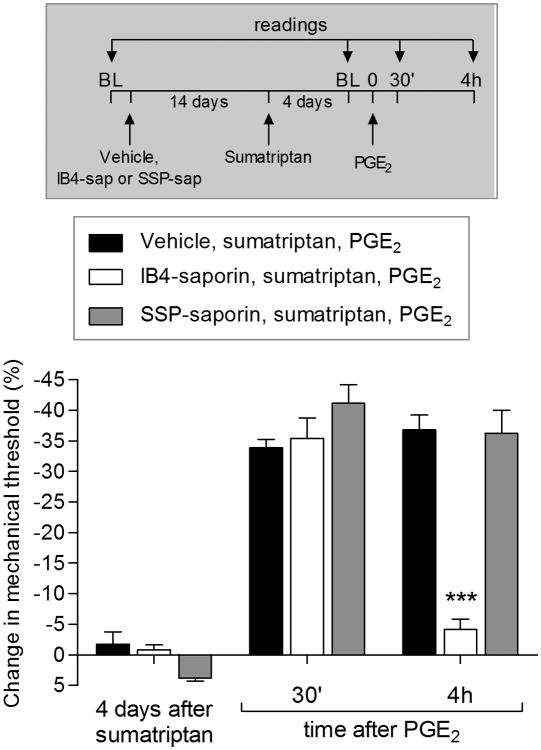
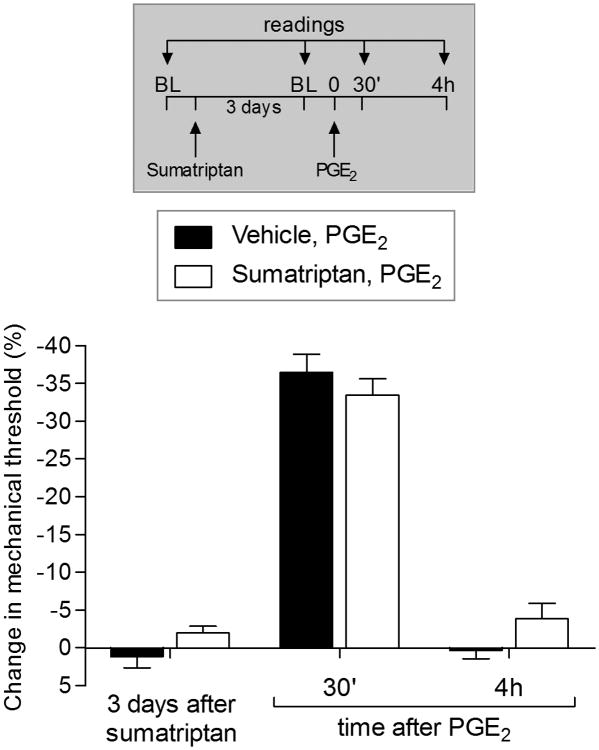
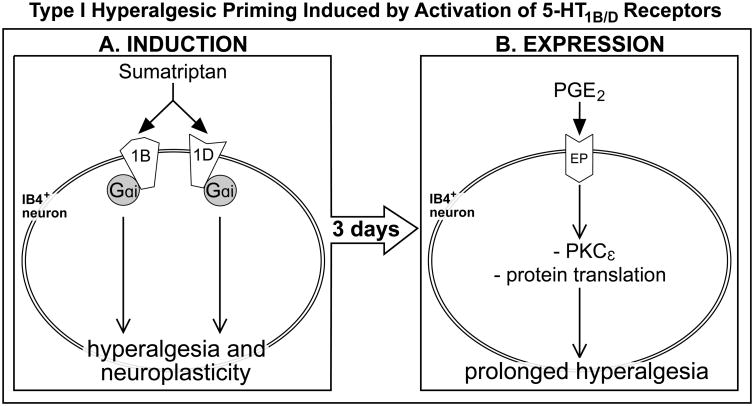
We have recently described a novel form of hyperalgesic priming (type II) induced by agonists at two clinically important Gi-protein-coupled receptors (Gi-GPCRs), mu-opioid and A1-adenosine. Like mu-opioids, the antimigraine triptans, which act at 5-HT1B/D Gi-GPCRs, have been implicated in pain chronification. We determined whether sumatriptan, a prototypical 5-HT1B/D agonist, produces type II priming. Characteristic of hyperalgesic priming, intradermal injection of sumatriptan (10 ng) induced a change in nociceptor function such that a subsequent injection of prostaglandin-E2 (PGE2) induces prolonged mechanical hyperalgesia. However, onset to priming was delayed 3 days, characteristic of type I priming. Also characteristic of type I priming, a protein kinase Cε, but not a protein kinase A inhibitor attenuated the prolongation phase of PGE2 hyperalgesia. The prolongation of PGE2 hyperalgesia was also permanently reversed by intradermal injection of cordycepin, a protein translation inhibitor. Also, hyperalgesic priming did not occur in animals pretreated with pertussis toxin or isolectin B4-positive nociceptor toxin, IB4-saporin. Finally, as observed for other agonists that induce type I priming, sumatriptan did not induce priming in female rats. The prolongation of PGE2 hyperalgesia induced by sumatriptan was partially prevented by coinjection of antagonists for the 5-HT1B and 5-HT1D, but not 5-HT7, serotonin receptors and completely prevented by coadministration of a combination of the 5-HT1B and 5-HT1D antagonists. Moreover, the injection of selective agonists, for 5-HT1B and 5-HT1D receptors, also induced hyperalgesic priming. Our results suggest that sumatriptan, which signals through Gi-GPCRs, induces type I hyperalgesic priming, unlike agonists at other Gi-GPCRs, which induce type II priming.
Figures
References
-
- Amrutkar DV, Ploug KB, Hay-Schmidt A, Porreca F, Olesen J, Jansen-Olesen I. mRNA expression of 5-hydroxytryptamine 1B, 1D, and 1F receptors and their role in controlling the release of calcitonin gene-related peptide in the rat trigeminovascular system. Pain. 2012;153(4):830–838. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
