

ERRγ Is Required for the Metabolic Maturation of Therapeutically Functional Glucose-Responsive β Cells
- PMID: 27076077
- PMCID: PMC4832237
- DOI: 10.1016/j.cmet.2016.03.005
ERRγ Is Required for the Metabolic Maturation of Therapeutically Functional Glucose-Responsive β Cells
Abstract
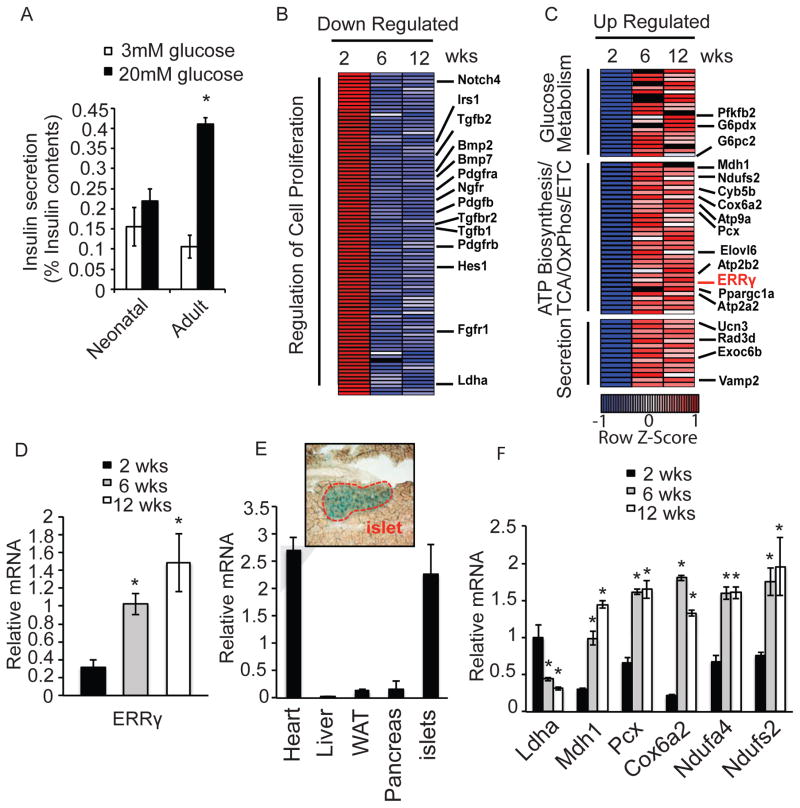
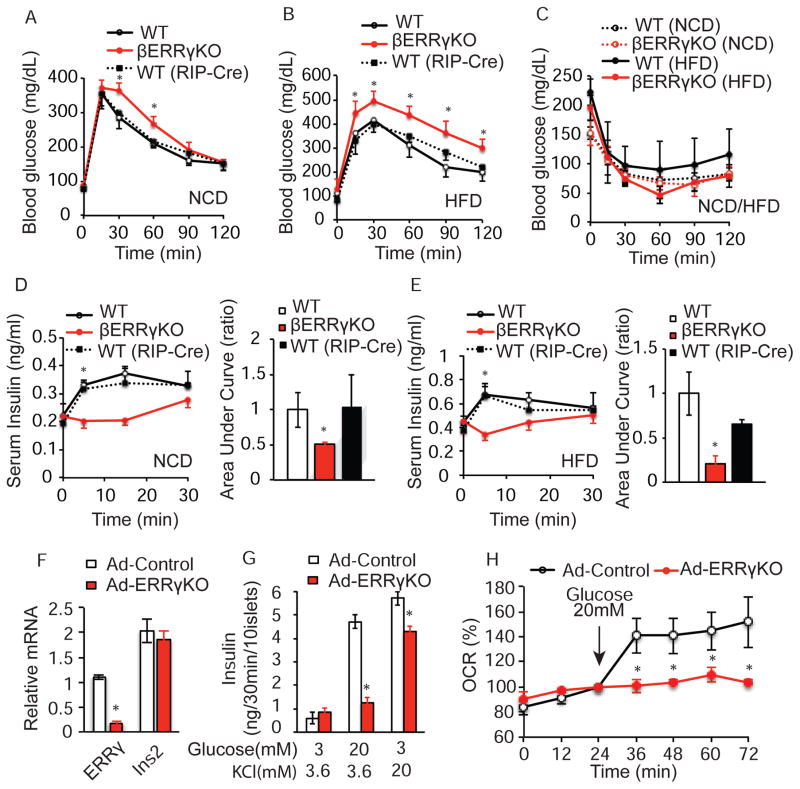
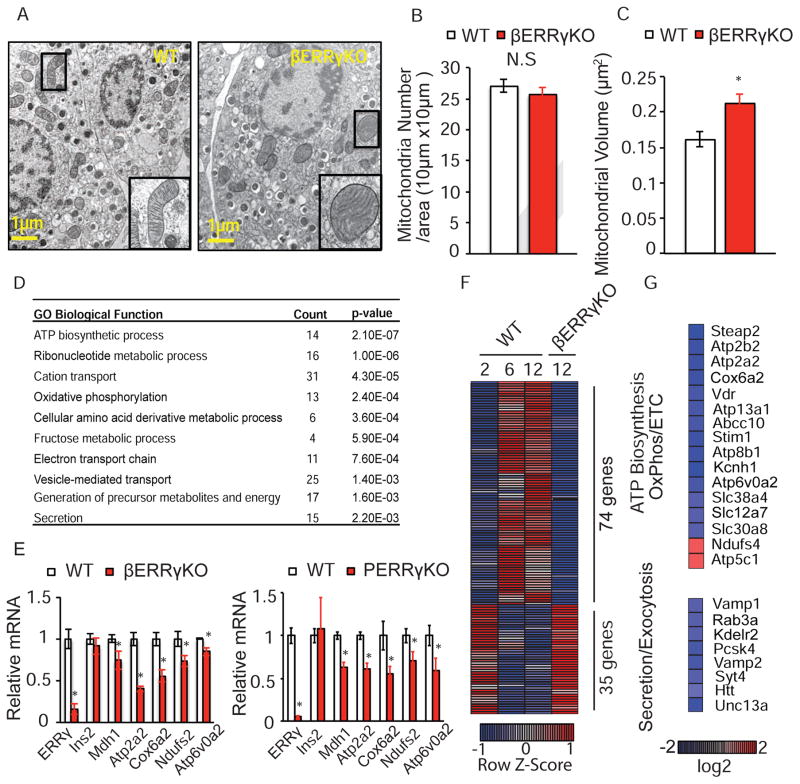
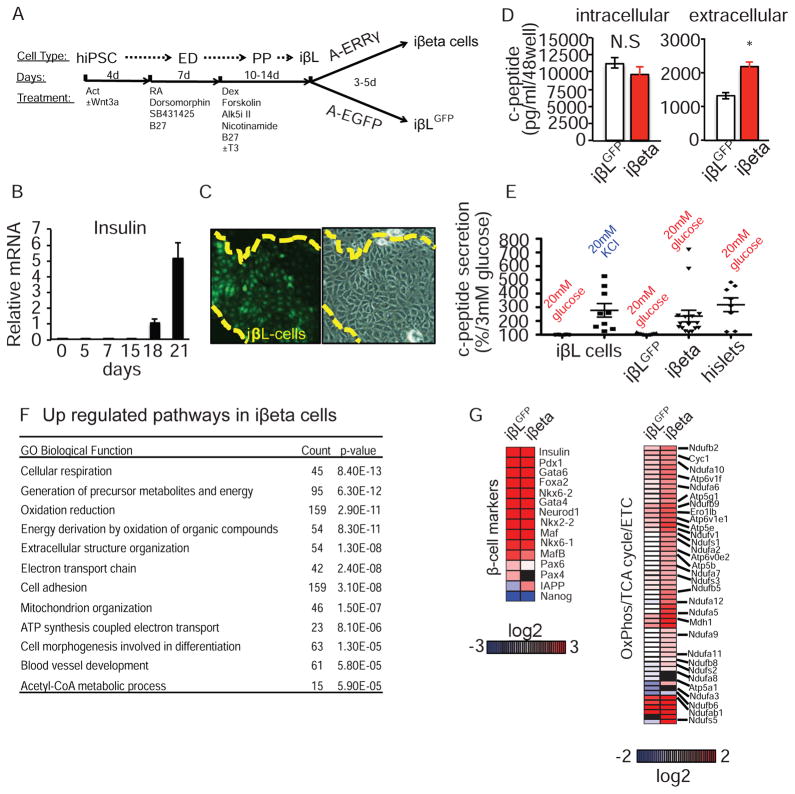
Pancreatic β cells undergo postnatal maturation to achieve maximal glucose-responsive insulin secretion, an energy intensive process. We identify estrogen-related receptor γ (ERRγ) expression as a hallmark of adult, but not neonatal β cells. Postnatal induction of ERRγ drives a transcriptional network activating mitochondrial oxidative phosphorylation, the electron transport chain, and ATP production needed to drive glucose-responsive insulin secretion. Mice deficient in β cell-specific ERRγ expression are glucose intolerant and fail to secrete insulin in response to a glucose challenge. Notably, forced expression of ERRγ in iPSC-derived β-like cells enables glucose-responsive secretion of human insulin in vitro, obviating in vivo maturation to achieve functionality. Moreover, these cells rapidly rescue diabetes when transplanted into β cell-deficient mice. These results identify a key role for ERRγ in β cell metabolic maturation, and offer a reproducible, quantifiable, and scalable approach for in vitro generation of functional human β cell therapeutics.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
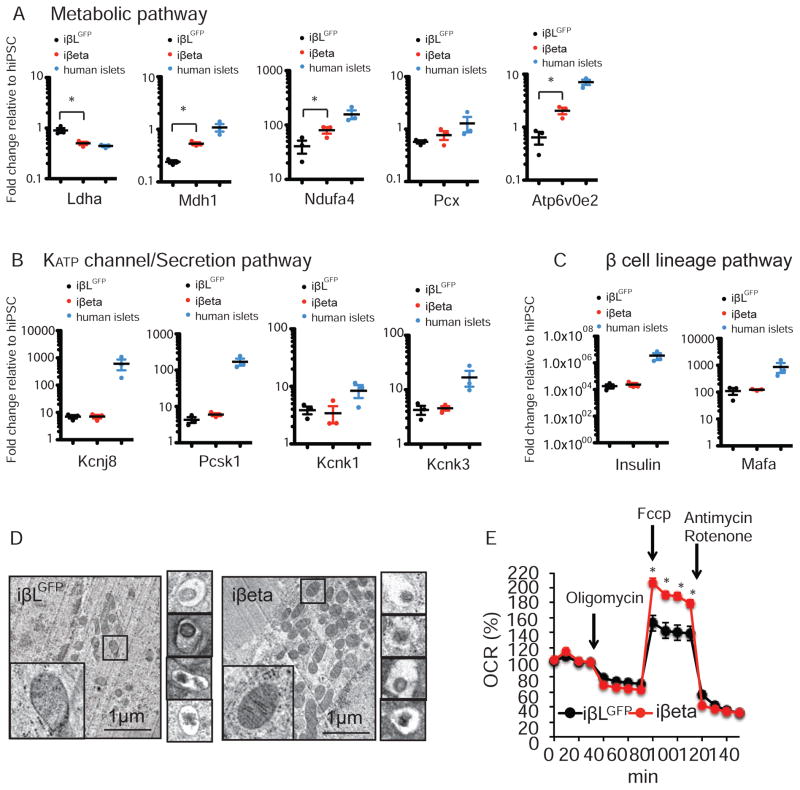
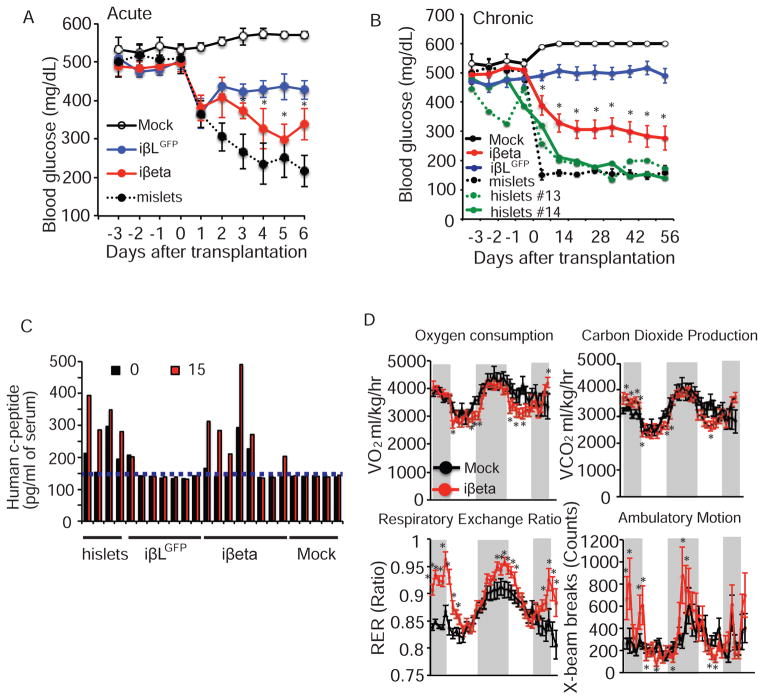
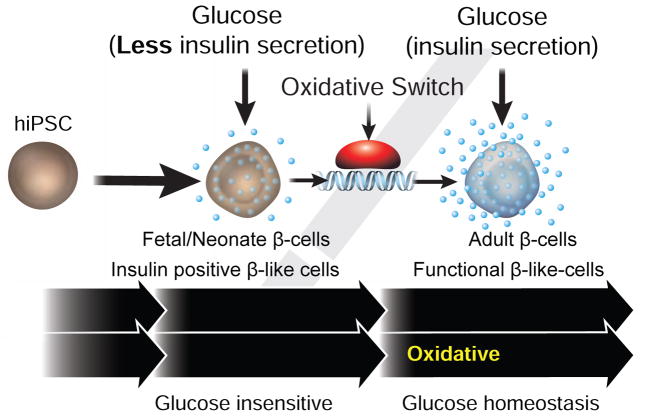
Comment in
-
Diabetes: New role for oestrogen-related receptor γ in β-cell maturation.Nat Rev Endocrinol. 2016 Jun;12(6):309. doi: 10.1038/nrendo.2016.61. Epub 2016 Apr 22. Nat Rev Endocrinol. 2016. PMID: 27109783 No abstract available.
-
ERRγ-A New Player in β Cell Maturation.Cell Metab. 2016 May 10;23(5):765-7. doi: 10.1016/j.cmet.2016.04.026. Cell Metab. 2016. PMID: 27166940
References
-
- Alaynick WA, Kondo RP, Xie W, He W, Dufour CR, Downes M, Jonker JW, Giles W, Naviaux RK, Giguere V, et al. ERRgamma directs and maintains the transition to oxidative metabolism in the postnatal heart. Cell metabolism. 2007;6:13–24. - PubMed
-
- Anello M, Lupi R, Spampinato D, Piro S, Masini M, Boggi U, Del Prato S, Rabuazzo AM, Purrello F, Marchetti P. Functional and morphological alterations of mitochondria in pancreatic beta cells from type 2 diabetic patients. Diabetologia. 2005;48:282–289. - PubMed
-
- Bliss CR, Sharp GW. Glucose-induced insulin release in islets of young rats: time-dependent potentiation and effects of 2-bromostearate. The American journal of physiology. 1992;263:E890–896. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P01 HL088093/HL/NHLBI NIH HHS/United States
- R01 HL105278/HL/NHLBI NIH HHS/United States
- HL088093/HL/NHLBI NIH HHS/United States
- P42 ES010337/ES/NIEHS NIH HHS/United States
- DK090962/DK/NIDDK NIH HHS/United States
- P30 DK063491/DK/NIDDK NIH HHS/United States
- ES010337/ES/NIEHS NIH HHS/United States
- R37 DK057978/DK/NIDDK NIH HHS/United States
- HL105278/HL/NHLBI NIH HHS/United States
- DK057978/DK/NIDDK NIH HHS/United States
- R24 DK090962/DK/NIDDK NIH HHS/United States
- R01 DK057978/DK/NIDDK NIH HHS/United States
- P30 CA014195/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
