

A Single Amino Acid Substitution in Poliovirus Nonstructural Protein 2CATPase Causes Conditional Defects in Encapsidation and Uncoating
- PMID: 27076638
- PMCID: PMC4936127
- DOI: 10.1128/JVI.02877-15
A Single Amino Acid Substitution in Poliovirus Nonstructural Protein 2CATPase Causes Conditional Defects in Encapsidation and Uncoating
Abstract
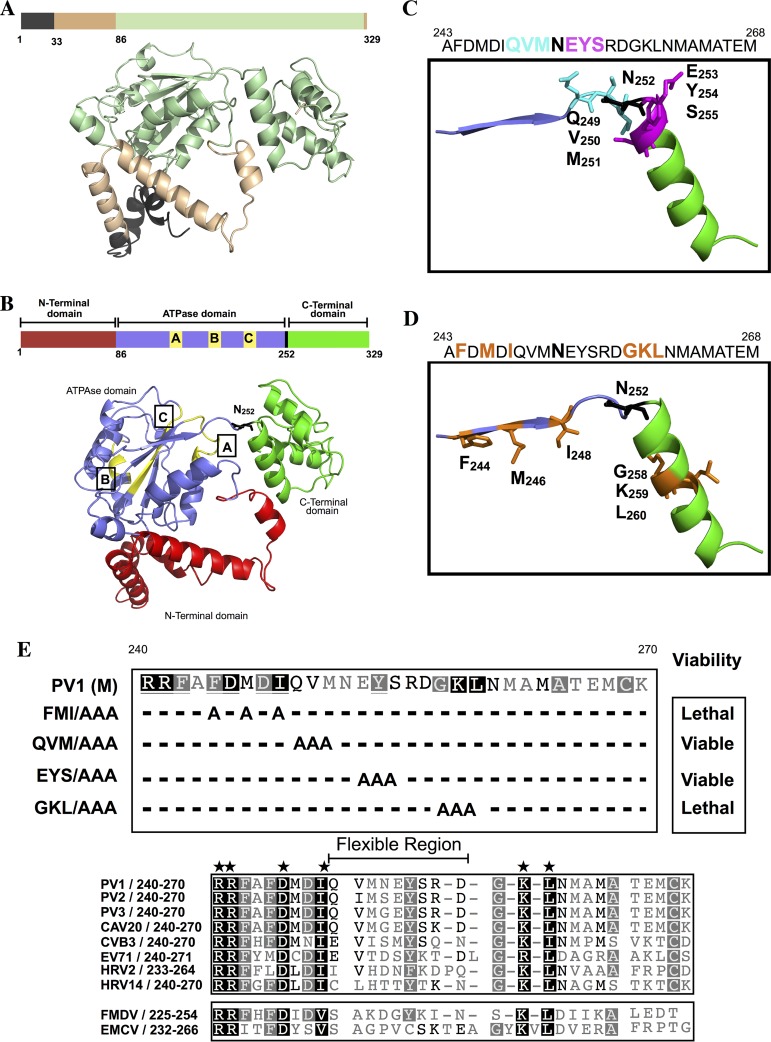
The specificity of encapsidation of C-cluster enteroviruses depends on an interaction between capsid proteins and nonstructural protein 2C(ATPase) In particular, residue N252 of poliovirus 2C(ATPase) interacts with VP3 of coxsackievirus A20, in the context of a chimeric virus. Poliovirus 2C(ATPase) has important roles both in RNA replication and encapsidation. In this study, we searched for additional sites in 2C(ATPase), near N252, that are required for encapsidation. Accordingly, segments adjacent to N252 were analyzed by combining triple and single alanine mutations to identify residues required for function. Two triple alanine mutants exhibited defects in RNA replication. The remaining two mutations, located in secondary structures in a predicted three-dimensional model of 2C(ATPase), caused lethal growth phenotypes. Most single alanine mutants, derived from the lethal variants, were either quasi-infectious and yielded variants with wild-type (wt) or temperature-sensitive (ts) growth phenotypes or had a lethal growth phenotype due to defective RNA replication. The K259A mutation, mapping to an α helix in the predicted structure of 2C(ATPase), resulted in a cold-sensitive virus. In vivo protein synthesis and virus production were strikingly delayed at 33°C relative to the wt, suggesting a defect in uncoating. Studies with a reporter virus indicated that this mutant is also defective in encapsidation at 33°C. Cell imaging confirmed a much-reduced production of K259A mature virus at 33°C relative to the wt. In conclusion, we have for the first time linked a cold-sensitive encapsidation defect in 2C(ATPase) (K259A) to a subsequent delay in uncoating of the virus particle at 33°C during the next cycle of infection.
Importance: Enterovirus morphogenesis, which involves the encapsidation of newly made virion RNA, is a process still poorly understood. Elucidation of this process is important for future drug development for a large variety of diseases caused by these agents. We have previously shown that the specificity of encapsidation of poliovirus and of C-cluster coxsackieviruses, which are prototypes of enteroviruses, is dependent on an interaction of capsid proteins with the multifunctional nonstructural protein 2C(ATPase) In this study, we have searched for residues in poliovirus 2C(ATPase), near a presumed capsid-interacting site, important for encapsidation. An unusual cold-sensitive mutant of 2C(ATPase) possessed a defect in encapsidation at 37°C and subsequently in uncoating during the next cycle of infection at 33°C. These studies not only reveal a new site in 2C(ATPase) that is involved in encapsidation but also identify a link between encapsidation and uncoating.
Copyright © 2016 Asare et al.
Figures


References
-
- Teterina NL, Kean KM, Gorbalenya AE, Agol VI, Girard M. 1992. Analysis of the functional significance of amino acid residues in the putative NTP-binding pattern of the poliovirus 2C protein. J Gen Virol 73:1977–1986. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
