

Simultaneous real-time measurement of EEG/EMG and L-glutamate in mice: A biosensor study of neuronal activity during sleep
- PMID: 27076812
- PMCID: PMC4827928
- DOI: 10.1016/j.jelechem.2010.12.031
Simultaneous real-time measurement of EEG/EMG and L-glutamate in mice: A biosensor study of neuronal activity during sleep
Abstract
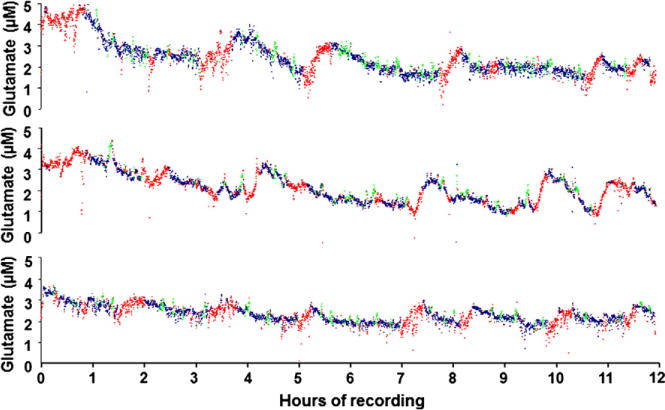
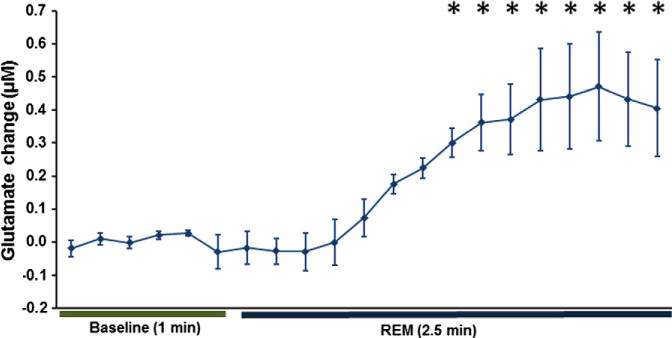
We report on electroencephalograph (EEG) and electromyograph (EMG) measurements concurrently with real-time changes in L-glutamate concentration. These data reveal a link between sleep state and extracellular neurotransmitter changes in a freely-moving (tethered) mouse. This study reveals, for the first time in mice, that the extracellular L-glutamate concentration in the pre-frontal cortex (PFC) increases during periods of extended wakefulness, decreases during extended sleep episodes and spikes during periods of REM sleep. Individual sleep epochs (10 s in duration) were scored as wake, slow-wave (SW) sleep or rapid eye movement (REM) sleep, and then correlated as a function of time with measured changes in L-glutamate concentrations. The observed L-glutamate levels show a statistically significant increase of 0.86 ± 0.26 μM (p < 0.05) over 37 wake episodes recorded from all mice (n = 6). Over the course of 49 measured sleep periods longer than 15 min, L-glutamate concentrations decline by a similar amount (0.88 ± 0.37 μM, p < 0.08). The analysis of 163 individual REM sleep episodes greater than one min in length across all mice (n = 6) demonstrates a significant rise in L-glutamate levels as compared to the 1 min preceding REM sleep onset (RM-ANOVA, DF = 20, F = 6.458, p < 0.001). The observed rapid changes in L-glutamate concentration during REM sleep last only between 1 and 3 min. The approach described can also be extended to other regions of the brain which are hypothesized to play a role in sleep. This study highlights the importance of obtaining simultaneous measurements of neurotransmitter levels in conjunction with sleep markers to help elucidate the underlying physiological and ultimately the genetic components of sleep.
Keywords: Continuous in vivo monitoring; Electroencephalography; Electromyography; Glutamate biosensor; Mouse sleep studies.
Figures






References
-
- Dement WC. History of sleep physiology and medicine. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practices of Sleep Medicine. fourth. W.B. Saunders Company; Philadelphia: 2005. pp. 1–12.
-
- Zepelin H, Siegel JM, Tobler I. Mammalian sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practices of Sleep Medicine. fourth. W.B. Saunders Company; Philadelphia: 2005. pp. 91–100.
-
- Steriade M. Brain electrical activity and sensory processing during waking and sleep states. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practices of Sleep Medicine. fourth. W.B. Saunders Company; Philadelphia: 2005. pp. 91–100.
-
- Carskadon MA, Dement WC. Normal human sleep: an overview. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practices of Sleep Medicine. W.B. Saunders; Philadelphia: 1994. pp. 889–913.
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous