

Reconstructing the demographic history of divergence between European river and brook lampreys using approximate Bayesian computations
- PMID: 27077007
- PMCID: PMC4830234
- DOI: 10.7717/peerj.1910
Reconstructing the demographic history of divergence between European river and brook lampreys using approximate Bayesian computations
Abstract
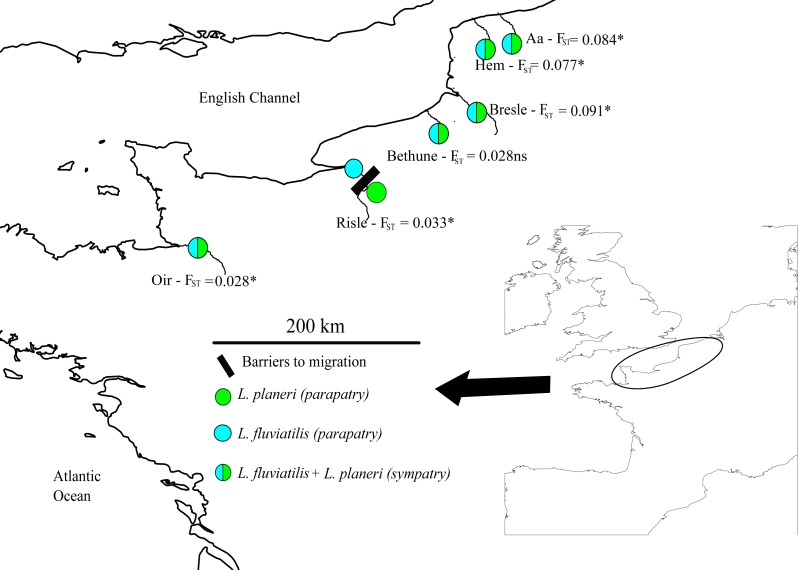
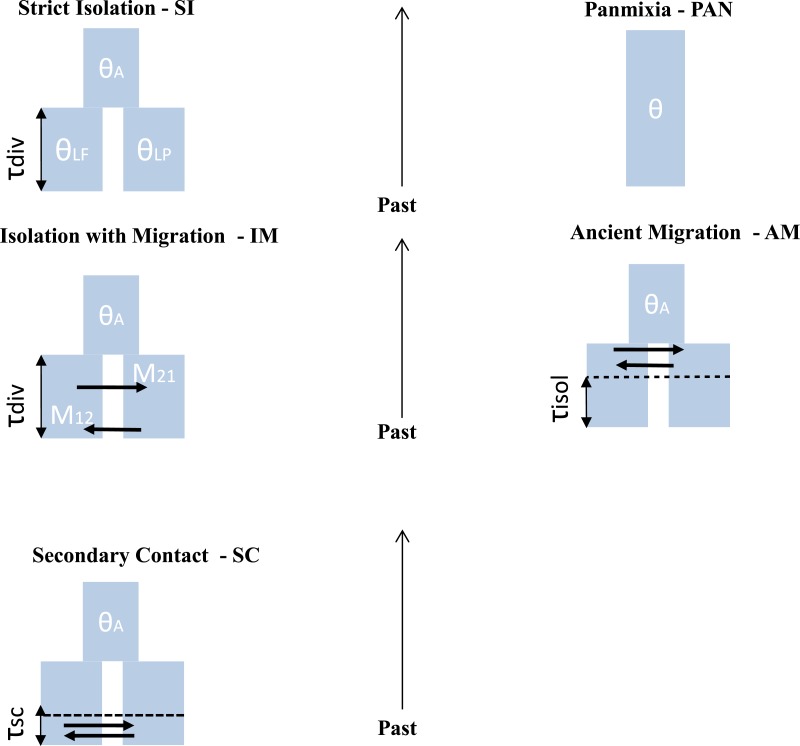
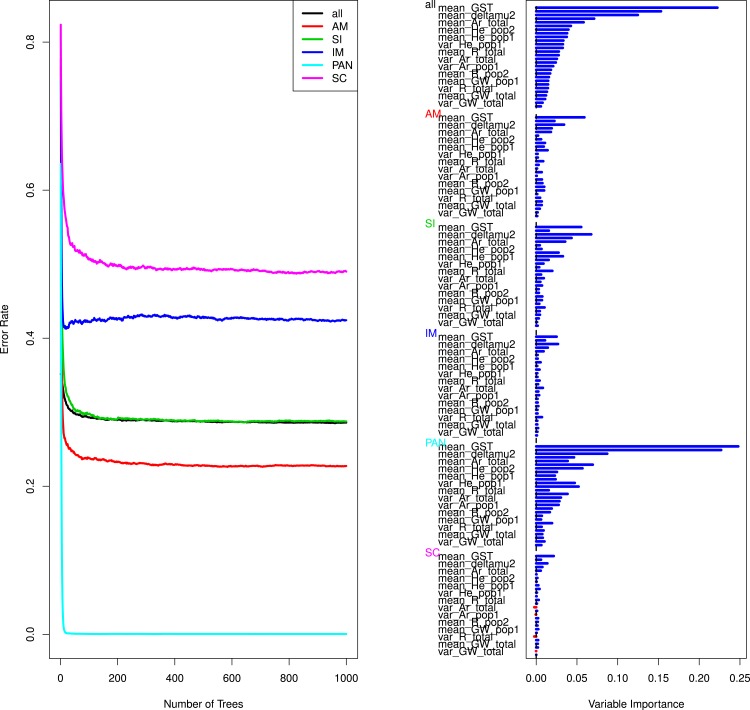
Inferring the history of isolation and gene flow during species divergence is a central question in evolutionary biology. The European river lamprey (Lampetra fluviatilis) and brook lamprey (L. planeri) show a low reproductive isolation but have highly distinct life histories, the former being parasitic-anadromous and the latter non-parasitic and freshwater resident. Here we used microsatellite data from six replicated population pairs to reconstruct their history of divergence using an approximate Bayesian computation framework combined with a random forest model. In most population pairs, scenarios of divergence with recent isolation were outcompeted by scenarios proposing ongoing gene flow, namely the Secondary Contact (SC) and Isolation with Migration (IM) models. The estimation of demographic parameters under the SC model indicated a time of secondary contact close to the time of speciation, explaining why SC and IM models could not be discriminated. In case of an ancient secondary contact, the historical signal of divergence is lost and neutral markers converge to the same equilibrium as under the less parameterized model allowing ongoing gene flow. Our results imply that models of secondary contacts should be systematically compared to models of divergence with gene flow; given the difficulty to discriminate among these models, we suggest that genome-wide data are needed to adequately reconstruct divergence history.
Keywords: Approximate Bayesian computation; Divergence history; Gene flow; Lampetra fluviatilis; Lampetra planeri; Random forest; Speciation.
Conflict of interest statement
The authors declare there are no competing interests.
Figures
References
-
- Barton NH, Hewitt GM. Analysis of hybrid zones. Annual Review of Ecology and Systematics. 1985;16:113–148. doi: 10.1146/annurev.es.16.110185.000553. - DOI
-
- Beamish RJ, Neville C-EM. The importance of size as an isolating mechanism in Lampreys. Copeia. 1992;1992:191–196. doi: 10.2307/1446551. - DOI
-
- Beaumont MA. Approximate Bayesian computation in evolution and ecology. Annual Review of Ecology, Evolution, and Systematics. 2010;41:379–406. doi: 10.1146/annurev-ecolsys-102209-144621. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
