

NLRX1 Sequesters STING to Negatively Regulate the Interferon Response, Thereby Facilitating the Replication of HIV-1 and DNA Viruses
- PMID: 27078069
- PMCID: PMC4833116
- DOI: 10.1016/j.chom.2016.03.001
NLRX1 Sequesters STING to Negatively Regulate the Interferon Response, Thereby Facilitating the Replication of HIV-1 and DNA Viruses
Abstract
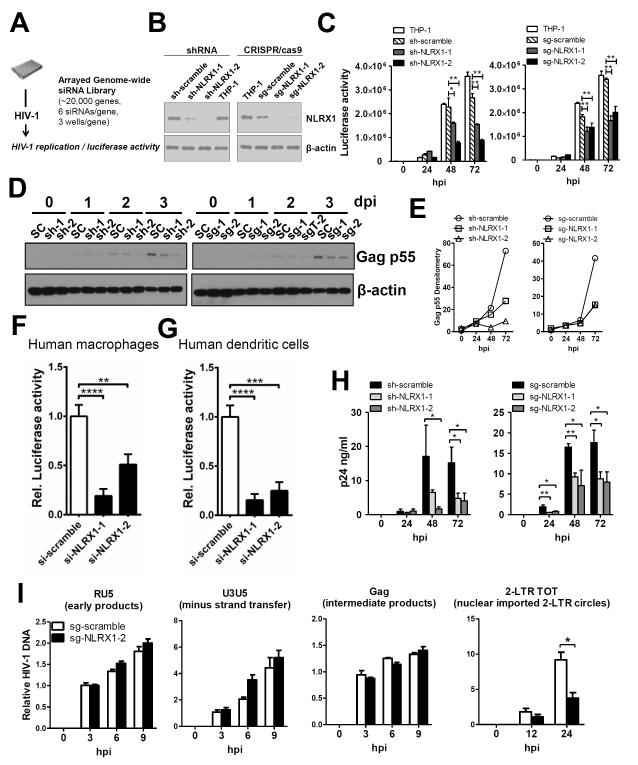
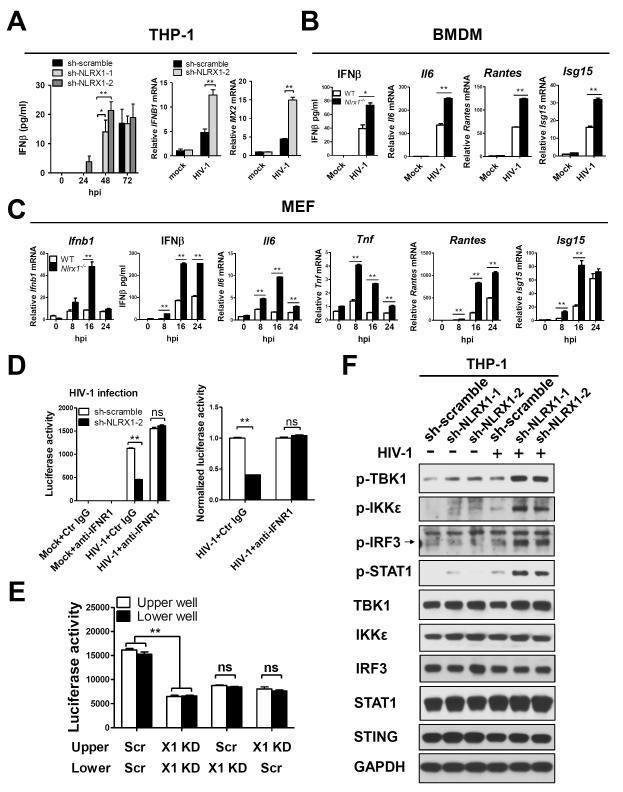
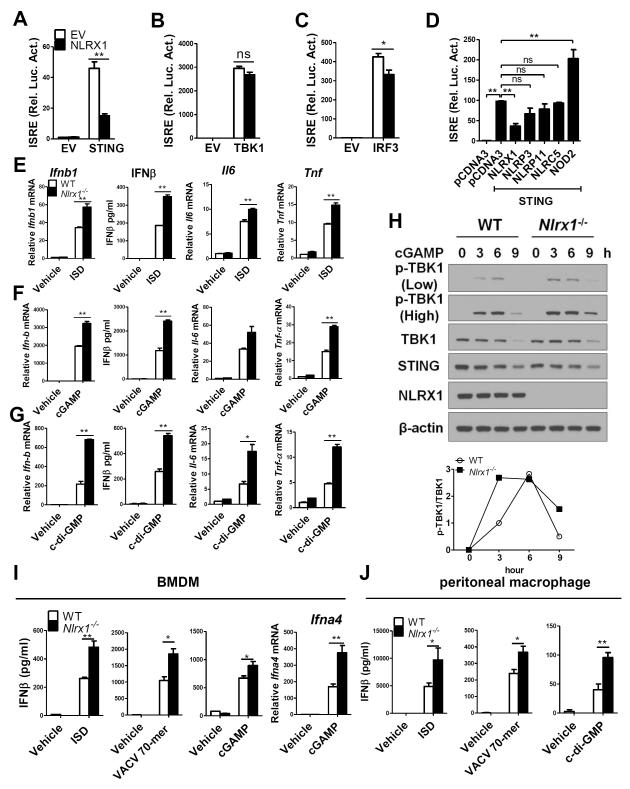
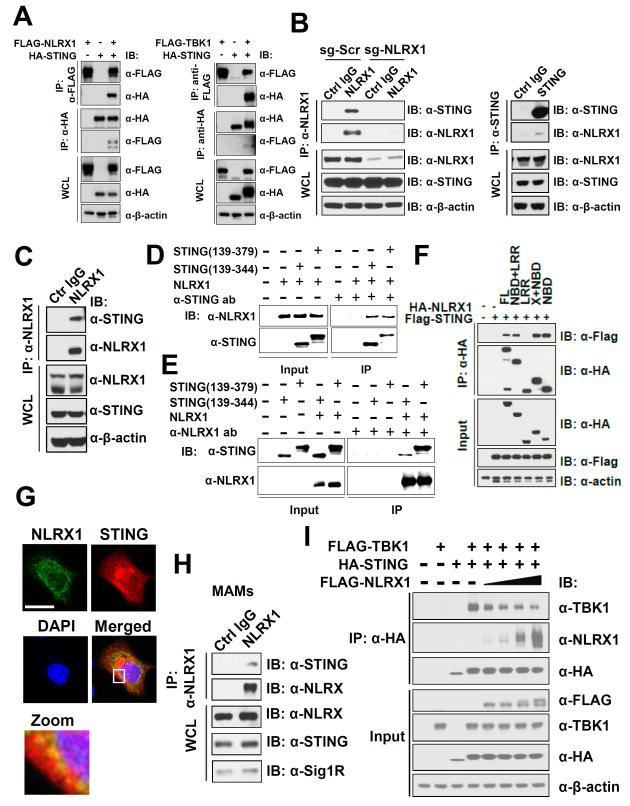
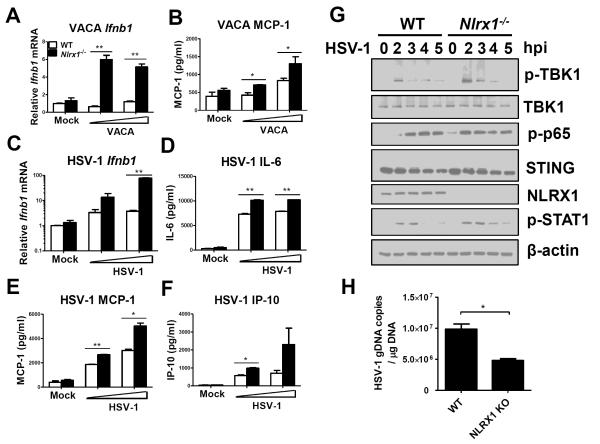
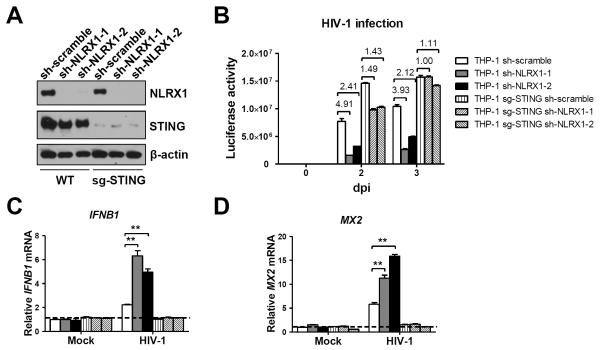
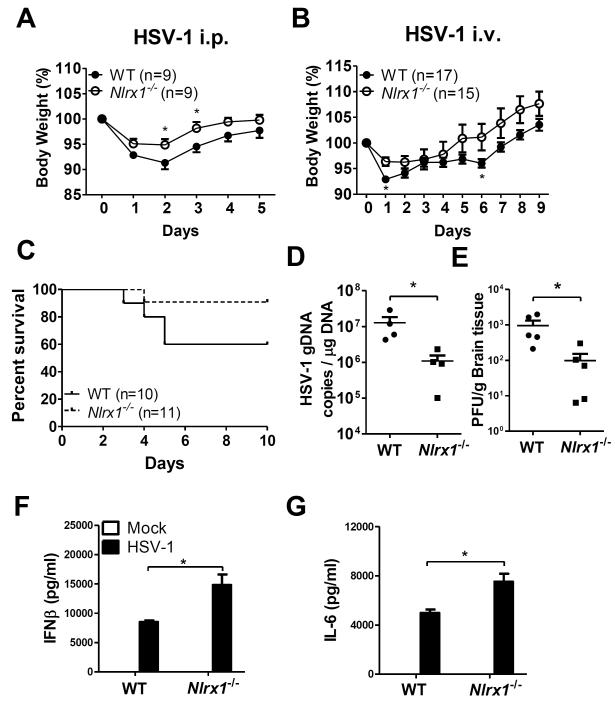
Understanding the negative regulators of antiviral immune responses will be critical for advancing immune-modulated antiviral strategies. NLRX1, an NLR protein that negatively regulates innate immunity, was previously identified in an unbiased siRNA screen as required for HIV infection. We find that NLRX1 depletion results in impaired nuclear import of HIV-1 DNA in human monocytic cells. Additionally, NLRX1 was observed to reduce type-I interferon (IFN-I) and cytokines in response to HIV-1 reverse-transcribed DNA. NLRX1 sequesters the DNA-sensing adaptor STING from interaction with TANK-binding kinase 1 (TBK1), which is a requisite for IFN-1 induction in response to DNA. NLRX1-deficient cells generate an amplified STING-dependent host response to cytosolic DNA, c-di-GMP, cGAMP, HIV-1, and DNA viruses. Accordingly, Nlrx1(-/-) mice infected with DNA viruses exhibit enhanced innate immunity and reduced viral load. Thus, NLRX1 is a negative regulator of the host innate immune response to HIV-1 and DNA viruses.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Abdul-Sater AA, Said-Sadier N, Lam VM, Singh B, Pettengill MA, Soares F, Tattoli I, Lipinski S, Girardin SE, Rosenstiel P, et al. Enhancement of reactive oxygen species production and chlamydial infection by the mitochondrial Nod-like family member NLRX1. The Journal of biological chemistry. 2010;285:41637–41645. - PMC - PubMed
-
- Altfeld M, Gale M., Jr. Innate immunity against HIV-1 infection. Nature immunology. 2015;16:554–562. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous