

INaP selective inhibition reverts precocious inter- and motorneurons hyperexcitability in the Sod1-G93R zebrafish ALS model
- PMID: 27079797
- PMCID: PMC4832213
- DOI: 10.1038/srep24515
INaP selective inhibition reverts precocious inter- and motorneurons hyperexcitability in the Sod1-G93R zebrafish ALS model
Abstract
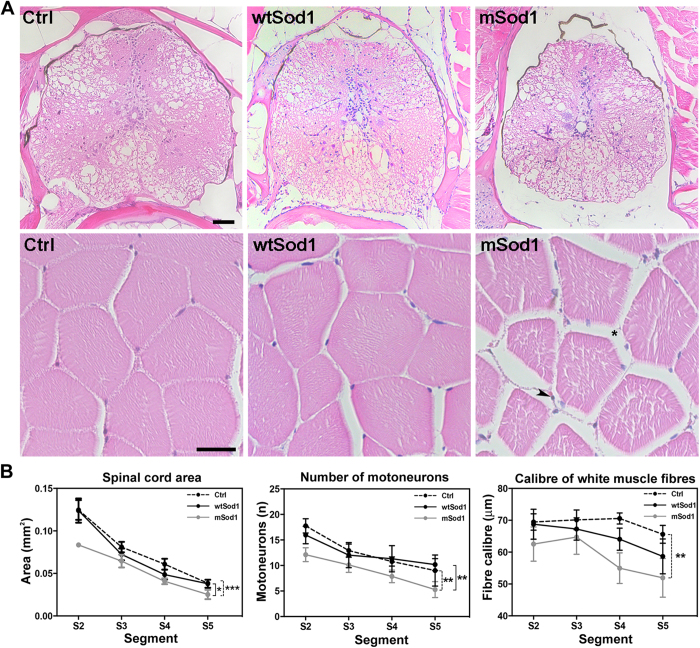
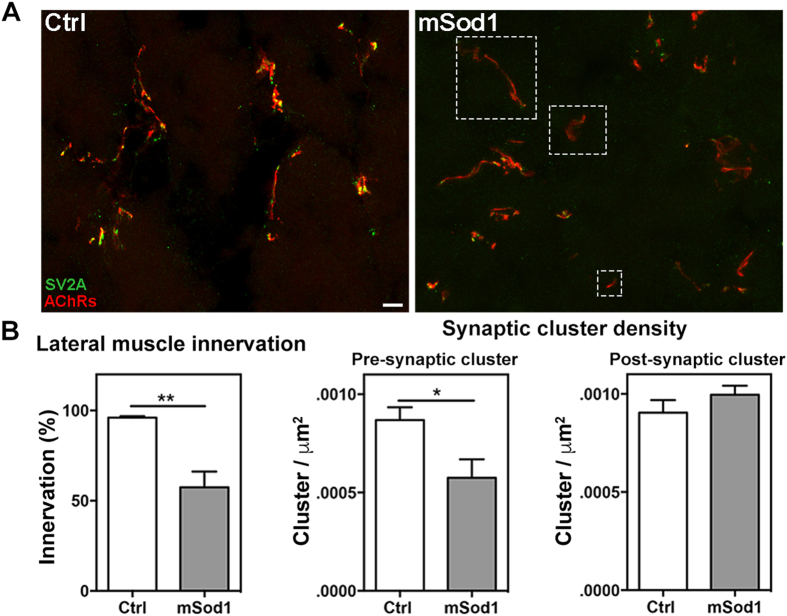
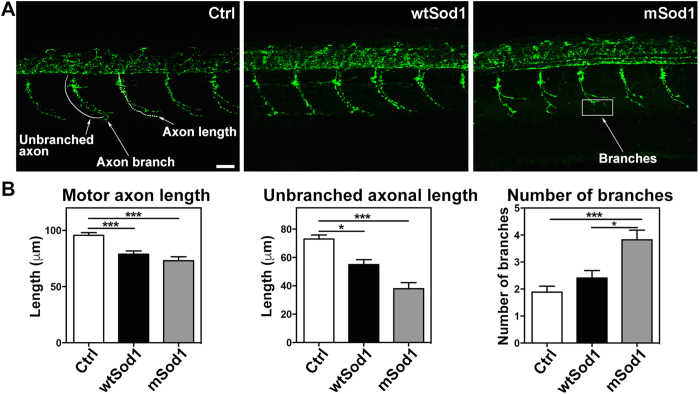
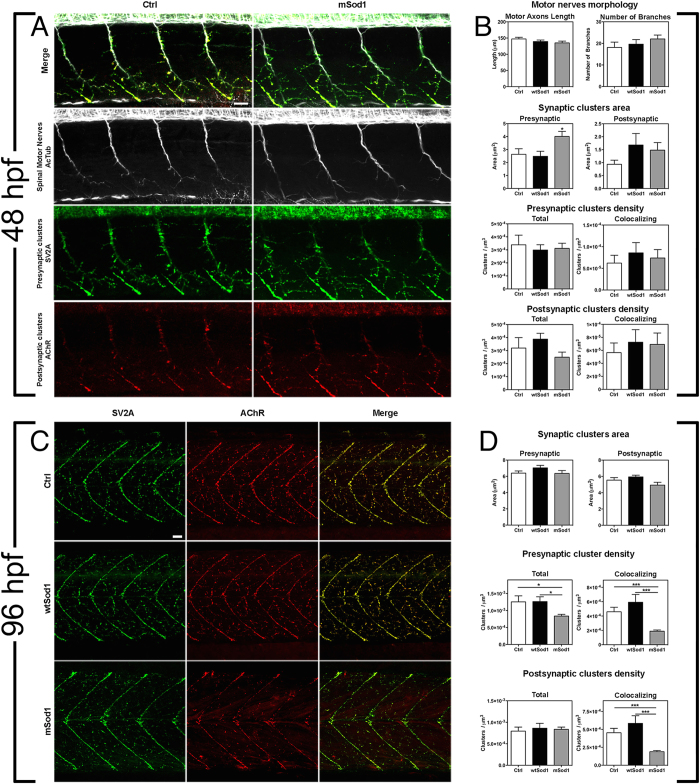
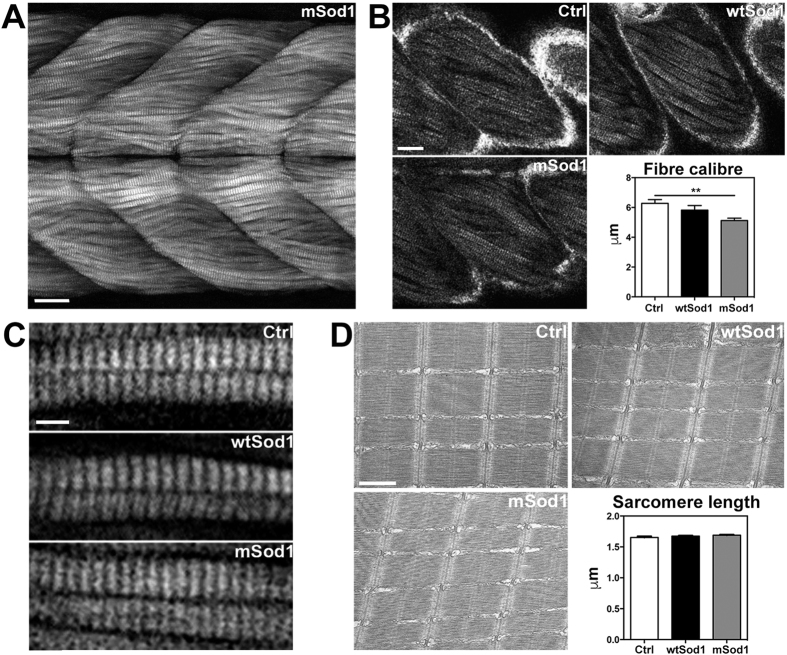
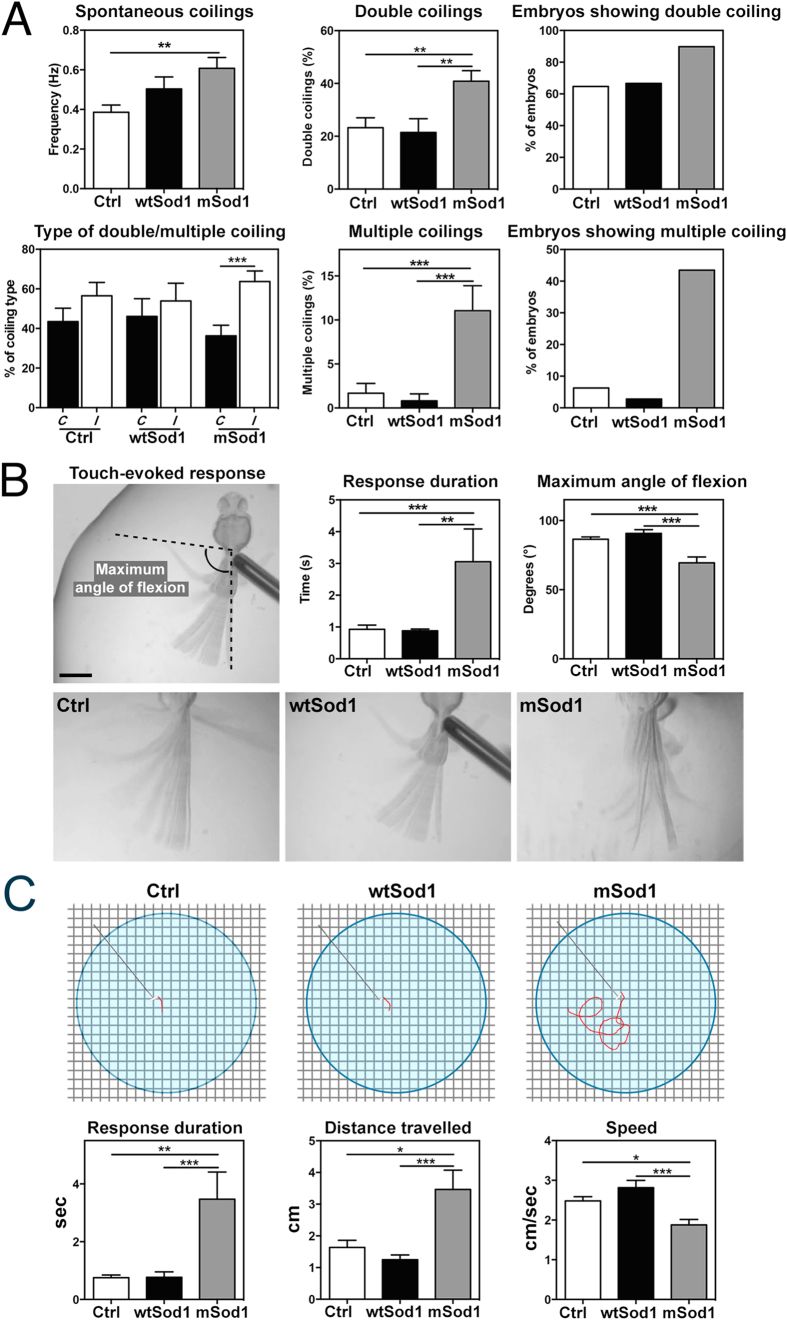
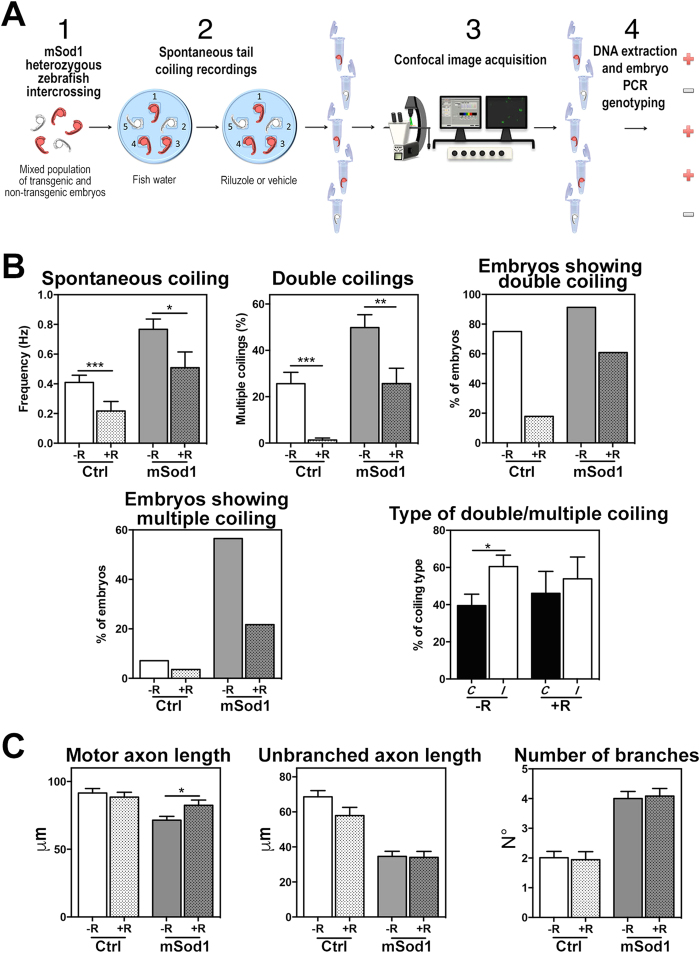
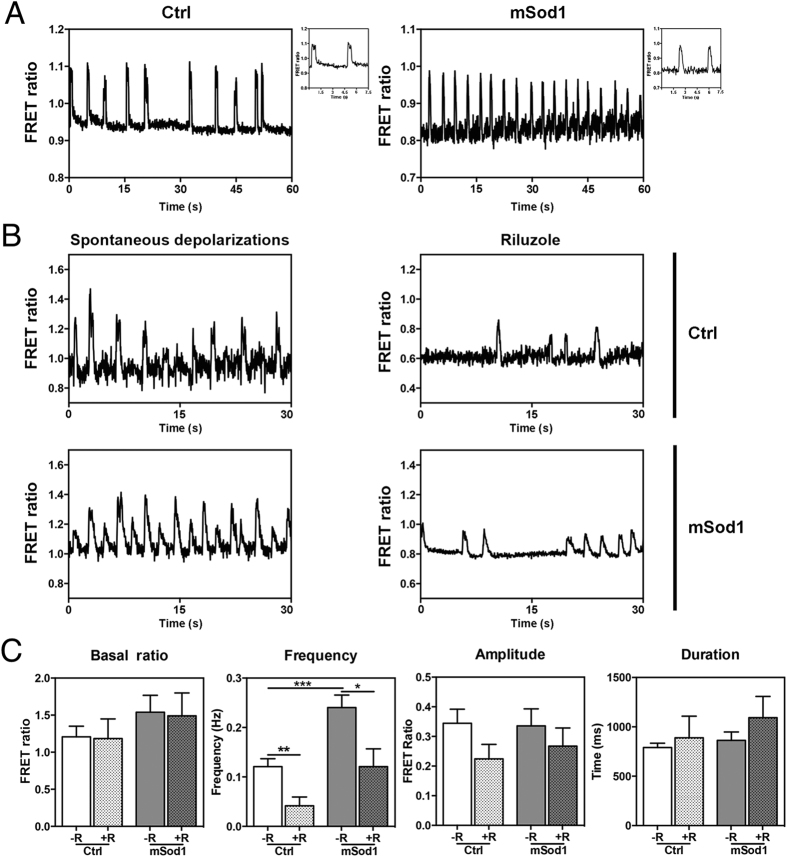
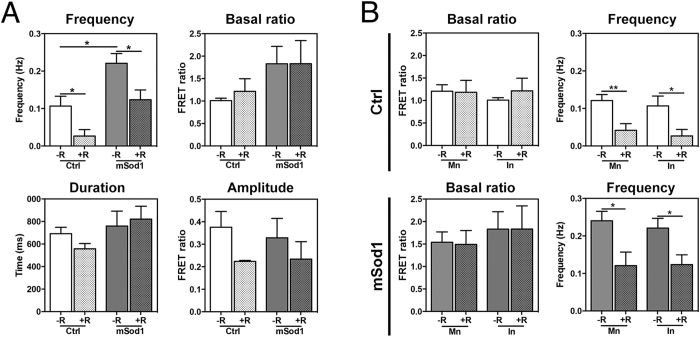
The pathogenic role of SOD1 mutations in amyotrophic lateral sclerosis (ALS) was investigated using a zebrafish disease model stably expressing the ALS-linked G93R mutation. In addition to the main pathological features of ALS shown by adult fish, we found remarkably precocious alterations in the development of motor nerve circuitry and embryo behavior, and suggest that these alterations are prompted by interneuron and motor neuron hyperexcitability triggered by anomalies in the persistent pacemaker sodium current INaP. The riluzole-induced modulation of INaP reduced spinal neuron excitability, reverted the behavioral phenotypes and improved the deficits in motor nerve circuitry development, thus shedding new light on the use of riluzole in the management of ALS. Our findings provide a valid phenotype-based tool for unbiased in vivo drug screening that can be used to develop new therapies.
Figures
References
-
- Vucic S., Rothstein J. D. & Kiernan M. C. Advances in treating amyotrophic lateral sclerosis: insights from pathophysiological studies. Trends Neurosci. 37, 433–442 (2014). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
