

Single-cell analysis of long non-coding RNAs in the developing human neocortex
- PMID: 27081004
- PMCID: PMC4831157
- DOI: 10.1186/s13059-016-0932-1
Single-cell analysis of long non-coding RNAs in the developing human neocortex
Abstract
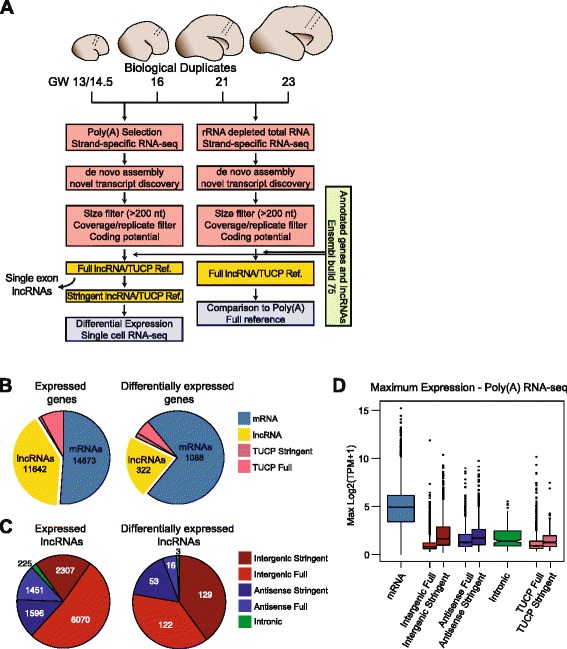
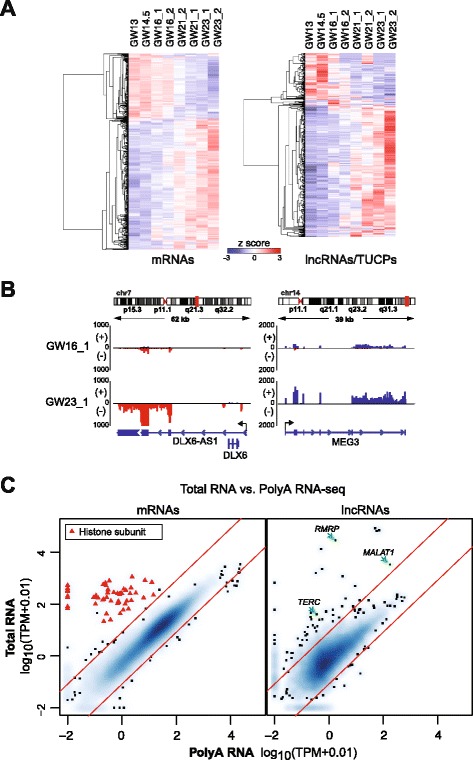
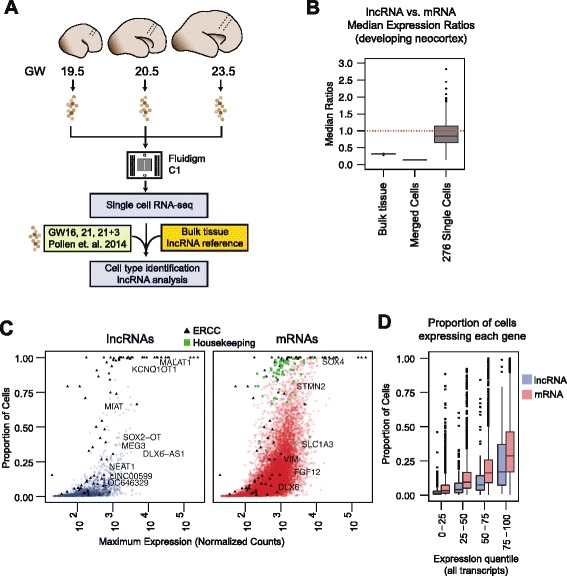
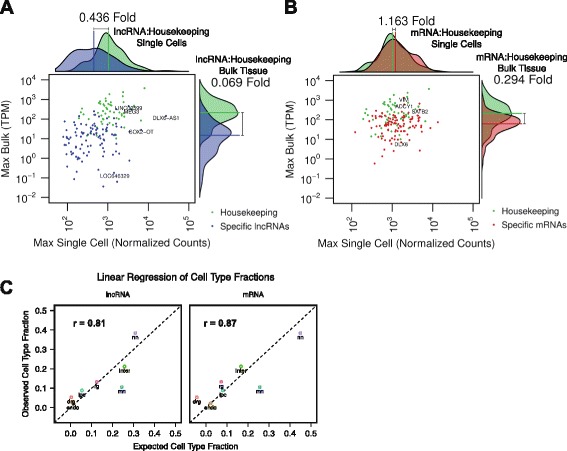
Background: Long non-coding RNAs (lncRNAs) comprise a diverse class of transcripts that can regulate molecular and cellular processes in brain development and disease. LncRNAs exhibit cell type- and tissue-specific expression, but little is known about the expression and function of lncRNAs in the developing human brain. Furthermore, it has been unclear whether lncRNAs are highly expressed in subsets of cells within tissues, despite appearing lowly expressed in bulk populations.
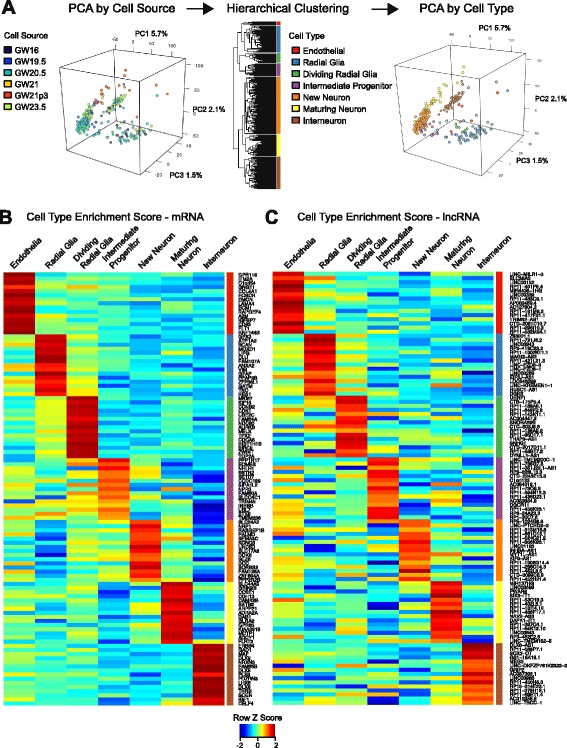
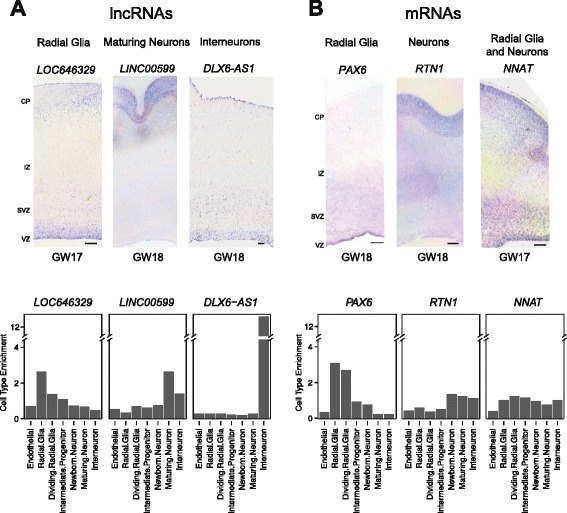
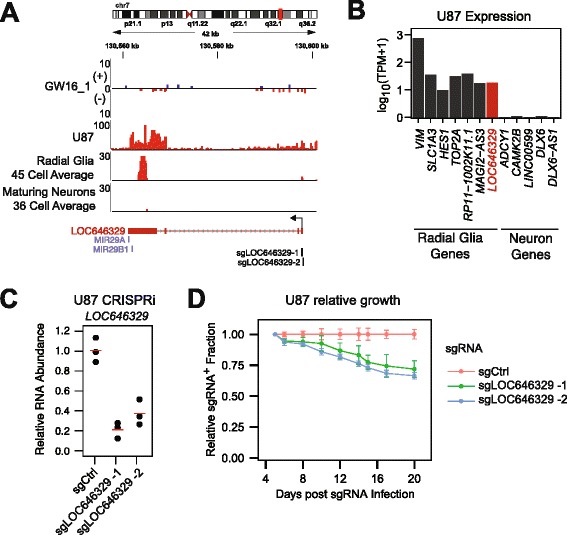
Results: We use strand-specific RNA-seq to deeply profile lncRNAs from polyadenylated and total RNA obtained from human neocortex at different stages of development, and we apply this reference to analyze the transcriptomes of single cells. While lncRNAs are generally detected at low levels in bulk tissues, single-cell transcriptomics of hundreds of neocortex cells reveal that many lncRNAs are abundantly expressed in individual cells and are cell type-specific. Notably, LOC646329 is a lncRNA enriched in single radial glia cells but is detected at low abundance in tissues. CRISPRi knockdown of LOC646329 indicates that this lncRNA regulates cell proliferation.
Conclusion: The discrete and abundant expression of lncRNAs among individual cells has important implications for both their biological function and utility for distinguishing neural cell types.
Keywords: CRISPRi; Developing brain; Single-cell RNA-seq; lncRNA.
Figures
Comment in
-
Single-cell profiling of lncRNAs in the developing human brain.Genome Biol. 2016 Apr 14;17:68. doi: 10.1186/s13059-016-0933-0. Genome Biol. 2016. PMID: 27079200 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
