

Comprehensively Characterizing the Thioredoxin Interactome In Vivo Highlights the Central Role Played by This Ubiquitous Oxidoreductase in Redox Control
- PMID: 27081212
- PMCID: PMC5083089
- DOI: 10.1074/mcp.M115.056440
Comprehensively Characterizing the Thioredoxin Interactome In Vivo Highlights the Central Role Played by This Ubiquitous Oxidoreductase in Redox Control
Abstract
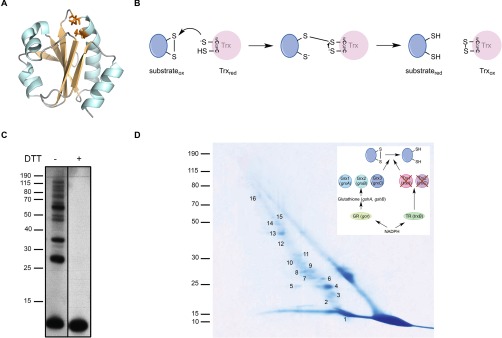
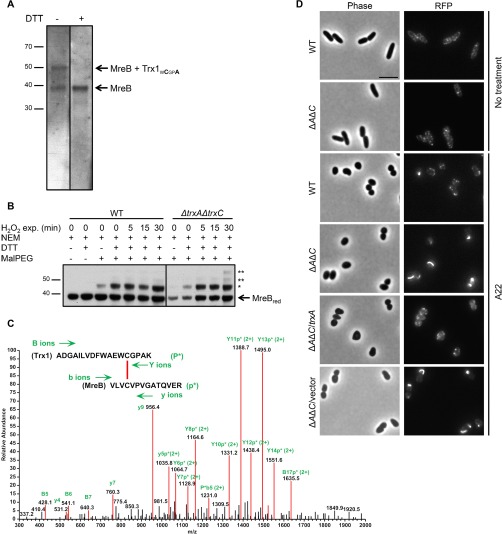
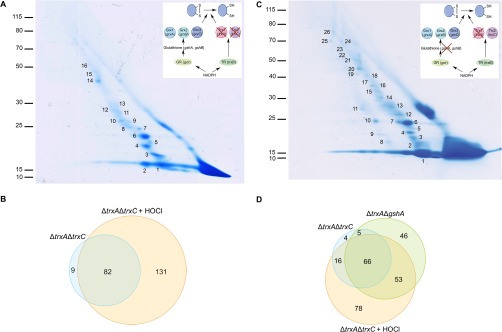
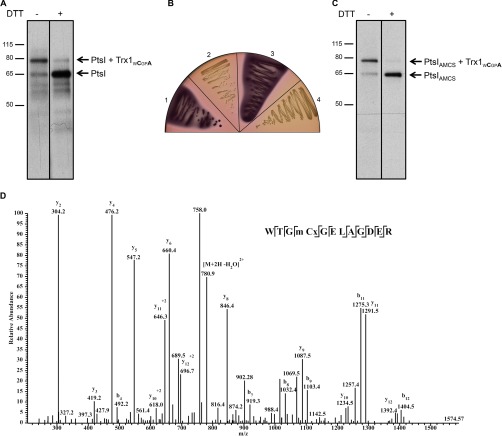
Thioredoxin (Trx) is a ubiquitous oxidoreductase maintaining protein-bound cysteine residues in the reduced thiol state. Here, we combined a well-established method to trap Trx substrates with the power of bacterial genetics to comprehensively characterize the in vivo Trx redox interactome in the model bacterium Escherichia coli Using strains engineered to optimize trapping, we report the identification of a total 268 Trx substrates, including 201 that had never been reported to depend on Trx for reduction. The newly identified Trx substrates are involved in a variety of cellular processes, ranging from energy metabolism to amino acid synthesis and transcription. The interaction between Trx and two of its newly identified substrates, a protein required for the import of most carbohydrates, PtsI, and the bacterial actin homolog MreB was studied in detail. We provide direct evidence that PtsI and MreB contain cysteine residues that are susceptible to oxidation and that participate in the formation of an intermolecular disulfide with Trx. By considerably expanding the number of Trx targets, our work highlights the role played by this major oxidoreductase in a variety of cellular processes. Moreover, as the dependence on Trx for reduction is often conserved across species, it also provides insightful information on the interactome of Trx in organisms other than E. coli.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Collet J. F., and Messens J. (2010) Structure, function, and mechanism of thioredoxin proteins. Antiox. Redox Signal. 13, 1205–1216 - PubMed
-
- Nakamura T., Nakamura H., Hoshino T., Ueda S., Wada H., and Yodoi J. (2005) Redox regulation of lung inflammation by thioredoxin. Antiox. Redox Signal. 7, 60–71 - PubMed
-
- Laurent T. C., Moore E. C., and Reichard P. (1964) Enzymatic synthesis of deoxyribonucleotides. IV. Isolation and characterization of thioredoxin, the hydrogen donor from Escherichia coli B. J. Biol. Chem. 239, 3436–3444 - PubMed
-
- Moore E. C., Reichard P., and Thelander L. (1964) Enzymatic synthesis of deoxyribonucleotides.V. Purification and properties of thioredoxin reductase from Escherichia coli B. J. Biol. Chem. 239, 3445–3452 - PubMed
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
