

Distinctive properties of metastasis-initiating cells
- PMID: 27083997
- PMCID: PMC4840296
- DOI: 10.1101/gad.277681.116
Distinctive properties of metastasis-initiating cells
Abstract
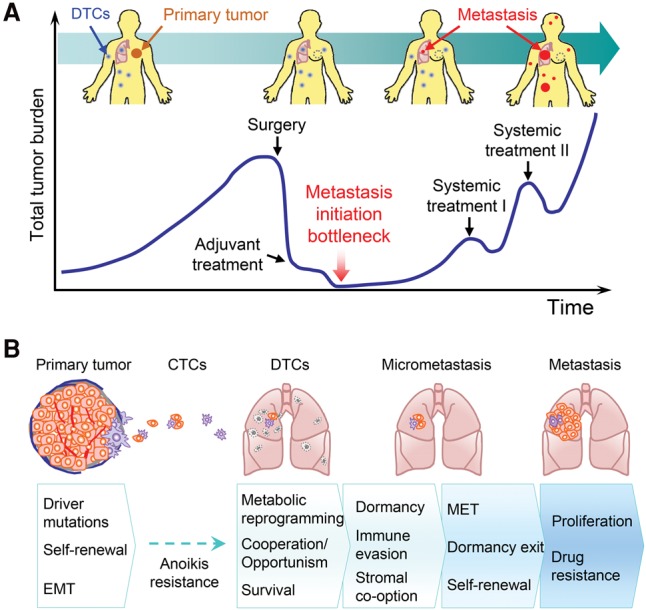
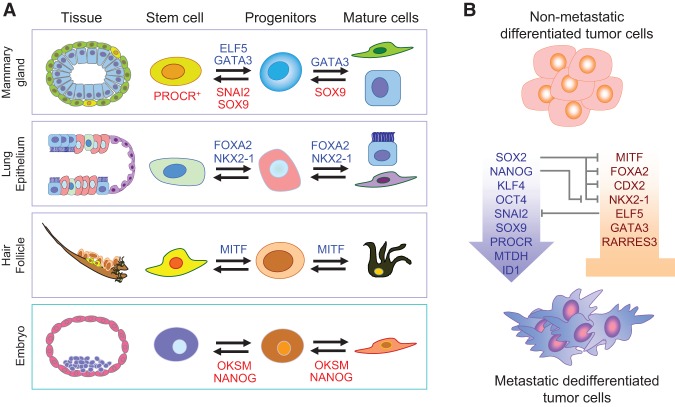
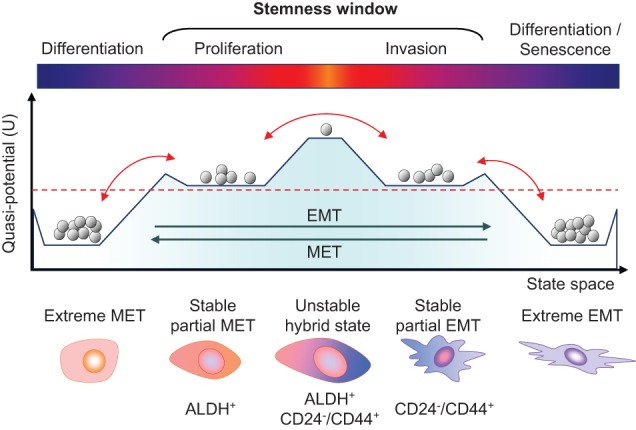
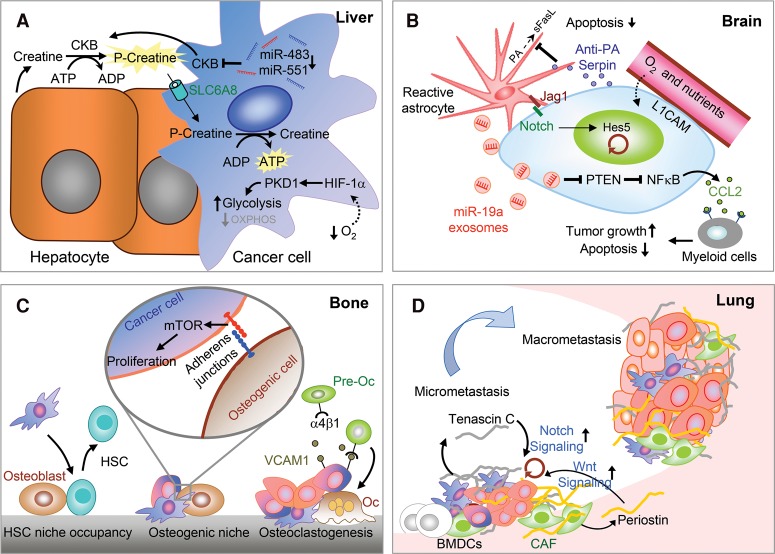
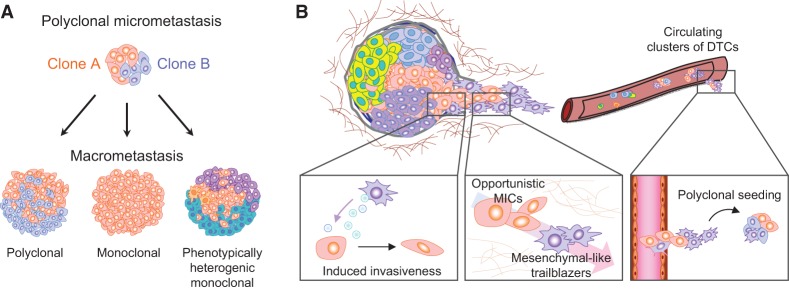
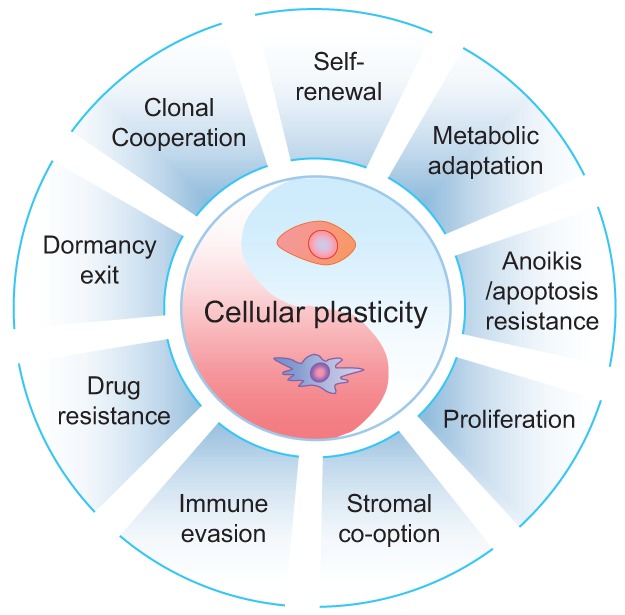
Primary tumors are known to constantly shed a large number of cancer cells into systemic dissemination, yet only a tiny fraction of these cells is capable of forming overt metastases. The tremendous rate of attrition during the process of metastasis implicates the existence of a rare and unique population of metastasis-initiating cells (MICs). MICs possess advantageous traits that may originate in the primary tumor but continue to evolve during dissemination and colonization, including cellular plasticity, metabolic reprogramming, the ability to enter and exit dormancy, resistance to apoptosis, immune evasion, and co-option of other tumor and stromal cells. Better understanding of the molecular and cellular hallmarks of MICs will facilitate the development and deployment of novel therapeutic strategies.
Keywords: cancer metastasis; epithelial–mesenchymal transition; metastasis-initiating cells; metastatic niche; plasticity.
© 2016 Celià-Terrassa and Kang; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Aguilar E, Marin de Mas I, Zodda E, Marin S, Morrish F, Selivanov V, Meca-Cortés O, Delowar H, Pons M, Izquierdo I, et al. 2016. Metabolic reprogramming and dependencies associated with epithelial cancer stem cells independent of the epithelial–mesenchymal transition program. Stem Cells 10.1002/stem.2286. - DOI - PMC - PubMed
-
- Barcellos-Hoff MH, Lyden D, Wang TC. 2013. The evolution of the cancer niche during multistage carcinogenesis. Nat Rev Cancer 13: 511–518. - PubMed
-
- Beck B, Lapouge G, Rorive S, Drogat B, Desaedelaere K, Delafaille S, Dubois C, Salmon I, Willekens K, Marine JC, et al. 2015. Different levels of Twist1 regulate skin tumor initiation, stemness, and progression. Cell Stem Cell 16: 67–79. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources