

The mammalian circadian clock and its entrainment by stress and exercise
- PMID: 27084533
- PMCID: PMC5138246
- DOI: 10.1007/s12576-016-0450-7
The mammalian circadian clock and its entrainment by stress and exercise
Abstract
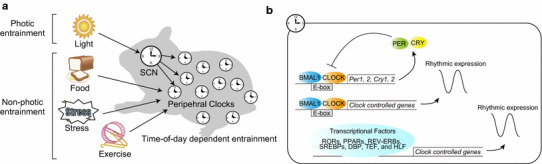
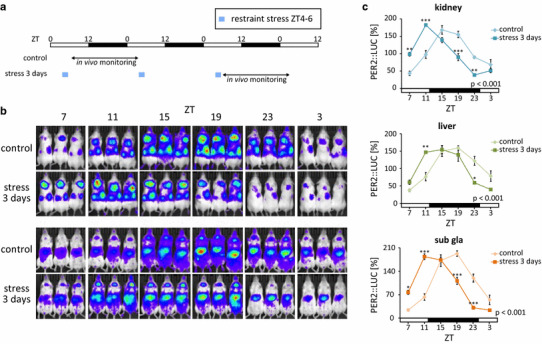
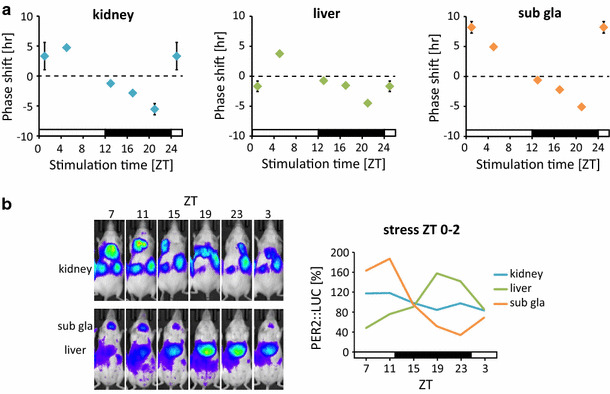
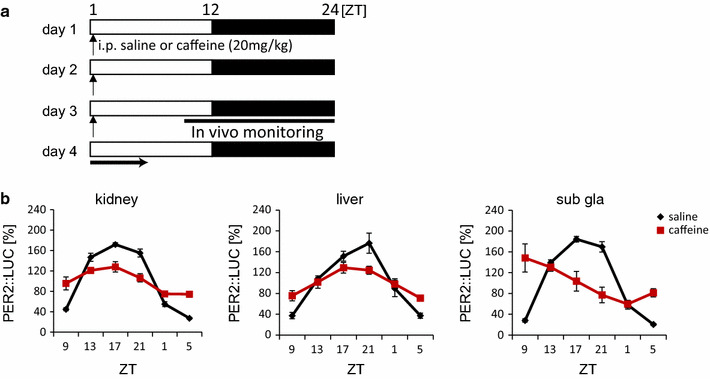
The mammalian circadian clock regulates day-night fluctuations in various physiological processes. The circadian clock consists of the central clock in the suprachiasmatic nucleus of the hypothalamus and peripheral clocks in peripheral tissues. External environmental cues, including light/dark cycles, food intake, stress, and exercise, provide important information for adjusting clock phases. This review focuses on stress and exercise as potent entrainment signals for both central and peripheral clocks, especially in regard to the timing of stimuli, types of stressors/exercises, and differences in the responses of rodents and humans. We suggest that the common signaling pathways of clock entrainment by stress and exercise involve sympathetic nervous activation and glucocorticoid release. Furthermore, we demonstrate that physiological responses to stress and exercise depend on time of day. Therefore, using exercise to maintain the circadian clock at an appropriate phase and amplitude might be effective for preventing obesity, diabetes, and cardiovascular disease.
Keywords: Liver; Mammalian circadian clock; Muscle; Oxidative stress.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Tahara Y, Kuroda H, Saito K, Nakajima Y, Kubo Y, Ohnishi N, Seo Y, Otsuka M, Fuse Y, Ohura Y, Komatsu T, Moriya Y, Okada S, Furutani N, Hirao A, Horikawa K, Kudo T, Shibata S. In vivo monitoring of peripheral circadian clocks in the mouse. Curr Biol. 2012;22(11):1029–1034. doi: 10.1016/j.cub.2012.04.009. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
