

T-cell selection in the thymus: a spatial and temporal perspective
- PMID: 27088910
- PMCID: PMC4938245
- DOI: 10.1111/imr.12398
T-cell selection in the thymus: a spatial and temporal perspective
Abstract
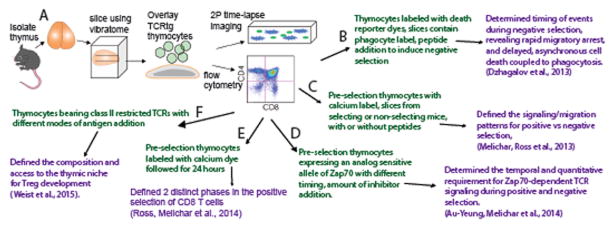
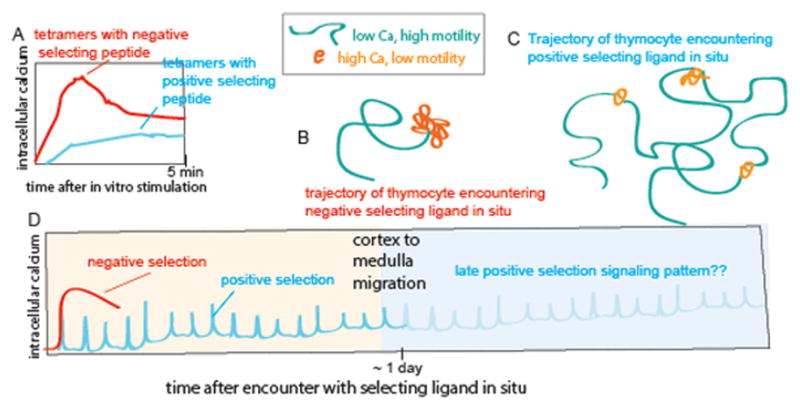
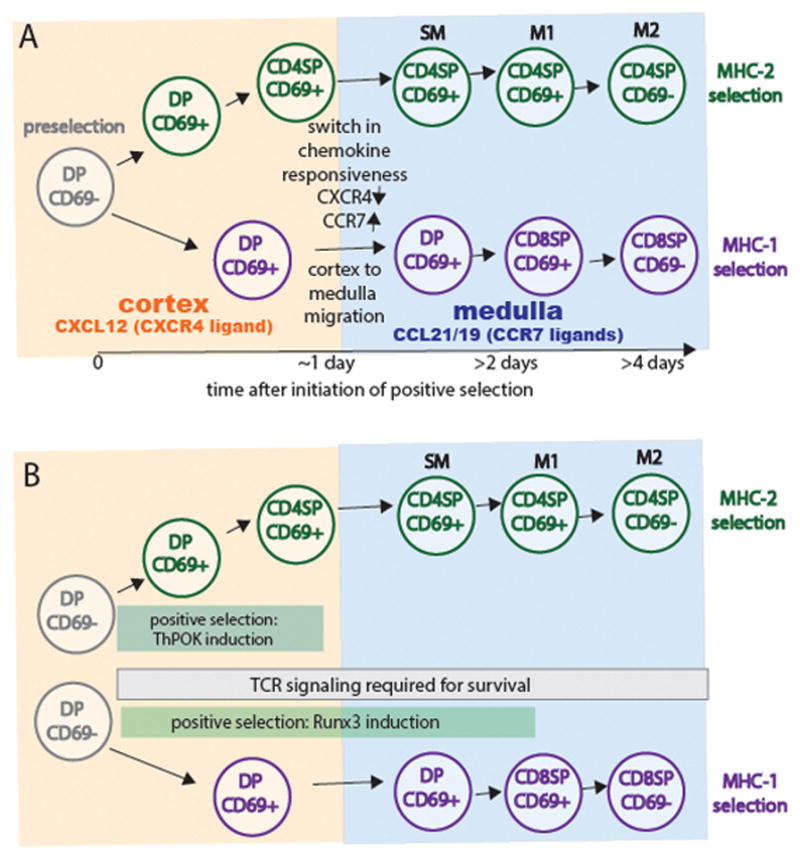
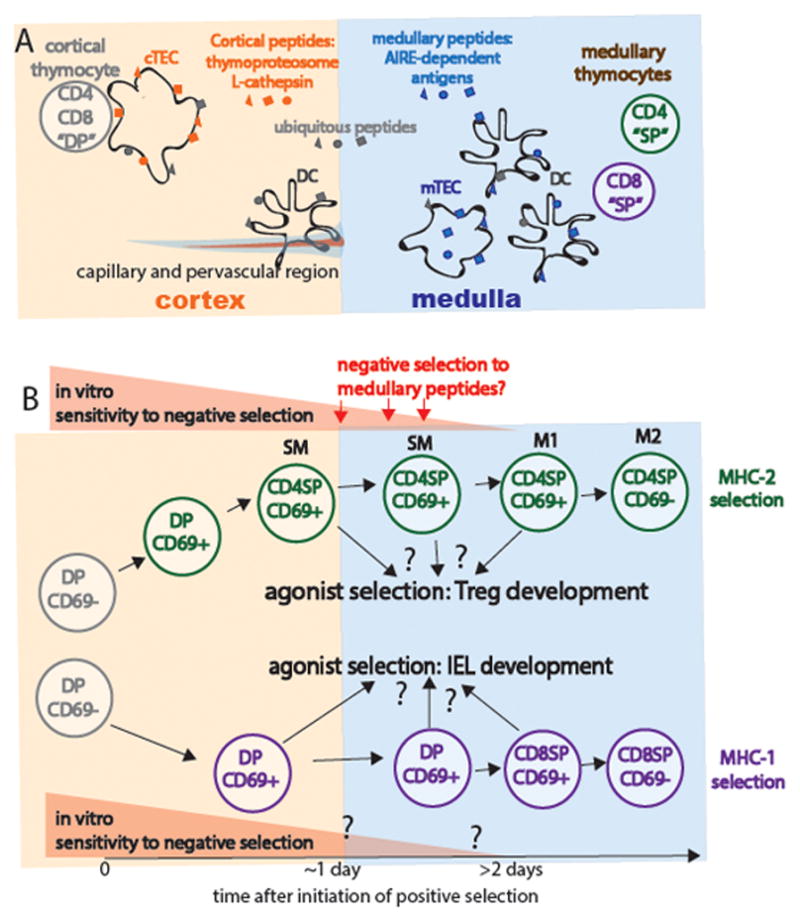
The ability of T cells to respond to a wide array of foreign antigens while avoiding reactivity to self is largely determined by cellular selection of developing T cells in the thymus. While a great deal is known about the cell types and molecules involved in T-cell selection in the thymus, our understanding of the spatial and temporal aspects of this process remain relatively poorly understood. Thymocytes are highly motile within the thymus and travel between specialized microenvironments at different phases of their development while interacting with distinct sets of self-peptides and peptide presenting cells. A knowledge of when, where, and how thymocytes encounter self-peptide MHC ligands at different stages of thymic development is key to understanding T-cell selection. In the past several years, our laboratory has investigated this topic using two-photon time-lapse microscopy to directly visualize thymocyte migration and signaling events, together with a living thymic slice preparation to provide a synchronized experimental model of T-cell selection in situ. Here, we discuss recent advances in our understanding of the temporal and spatial aspects of T-cell selection, highlighting our own work, and placing them in the context of work from other groups.
Keywords: T-cell antigen receptor; cell differentiation; chemokines; lineage commitment/specification; thymus.
© 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Ladi E, Yin X, Chtanova T, Robey EA. Thymic microenvironments for T cell differentiation and selection. Nat Immunol. 2006;7:338–343. - PubMed
-
- Takahama Y. Journey through the thymus: stromal guides for T-cell development and selection. Nat Rev Immunol. 2006;6:127–135. - PubMed
-
- Starr TK, Jameson SC, Hogquist KA. Positive and negative selection of T cells. Annu Rev Immunol. 2003;21:139–176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
