

Peyer's patches: organizing B-cell responses at the intestinal frontier
- PMID: 27088918
- PMCID: PMC4835804
- DOI: 10.1111/imr.12400
Peyer's patches: organizing B-cell responses at the intestinal frontier
Abstract
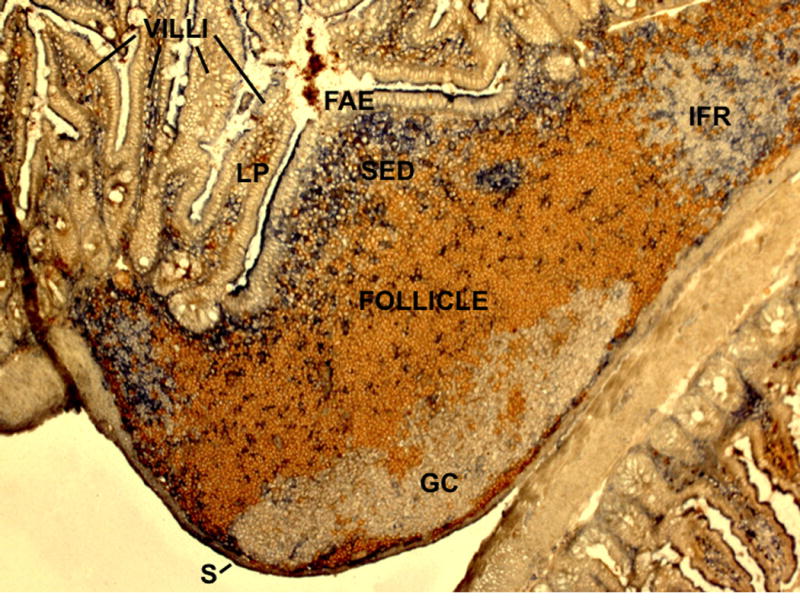
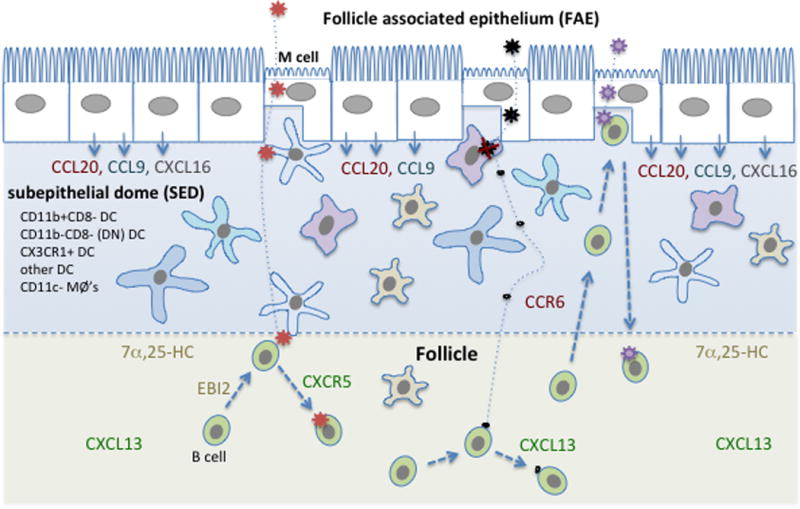
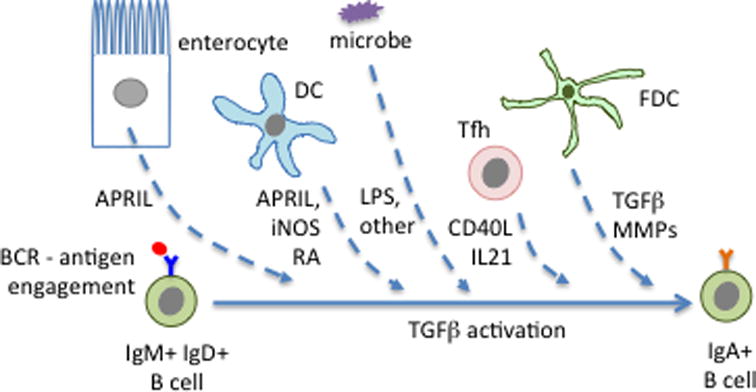
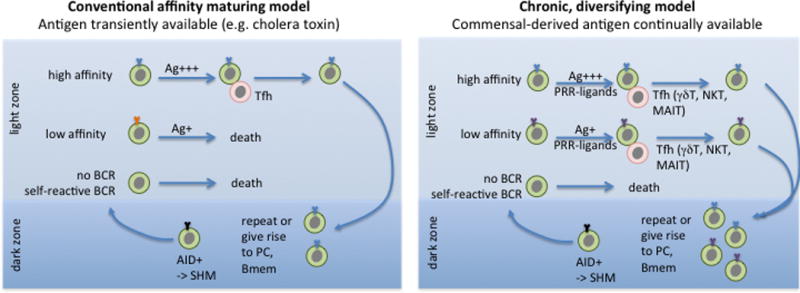
Secondary lymphoid tissues share the important function of bringing together antigens and rare antigen-specific lymphocytes to foster induction of adaptive immune responses. Peyer's patches (PPs) are unique compared to other secondary lymphoid tissues in their continual exposure to an enormous diversity of microbiome- and food-derived antigens and in the types of pathogens they encounter. Antigens are delivered to PPs by specialized microfold (M) epithelial cells and they may be captured and presented by resident dendritic cells (DCs). In accord with their state of chronic microbial antigen exposure, PPs exhibit continual germinal center (GC) activity. These GCs not only contribute to the generation of B cells and plasma cells producing somatically mutated gut antigen-specific IgA antibodies but have also been suggested to support non-specific antigen diversification of the B-cell repertoire. Here, we review current understanding of how PPs foster B-cell encounters with antigen, how they favor isotype switching to the secretory IgA isotype, and how their GC responses may uniquely contribute to mucosal immunity.
Keywords: B cells; antibodies; cell trafficking; dendritic cells; mucosa; repertoire development.
© 2016 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare they have no financial conflict of interest
Figures
References
-
- Brandtzaeg P. Role of secretory antibodies in the defence against infections. Int J Med Microbiol. 2003;293:3–15. - PubMed
-
- Macpherson AJ, Koller Y, McCoy KD. The bilateral responsiveness between intestinal microbes and IgA. Trends Immunol. 2015;36:460–470. - PubMed
-
- Fagarasan S, Kawamoto S, Kanagawa O, Suzuki K. Adaptive immune regulation in the gut: T cell-dependent and T cell-independent IgA synthesis. Annu Rev Immunol. 2010;28:243–273. - PubMed
-
- Macpherson AJ, Geuking MB, Slack E, Hapfelmeier S, McCoy KD. The habitat, double life, citizenship, and forgetfulness of IgA. Immulogical Reviews. 2011;245:132–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
