

Recurrent activating mutations of G-protein-coupled receptor CYSLTR2 in uveal melanoma
- PMID: 27089179
- PMCID: PMC5032652
- DOI: 10.1038/ng.3549
Recurrent activating mutations of G-protein-coupled receptor CYSLTR2 in uveal melanoma
Abstract
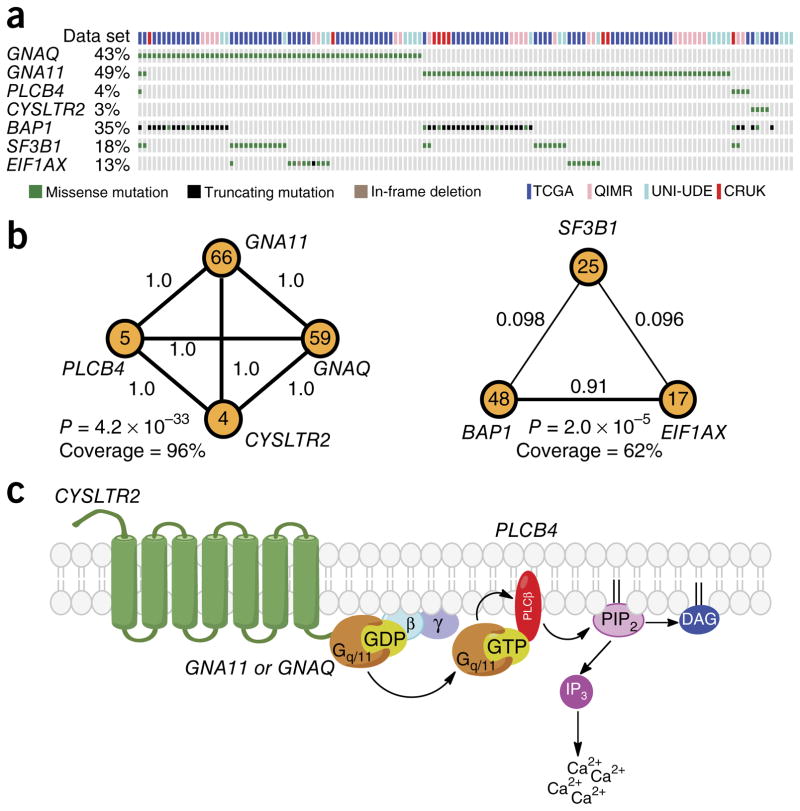
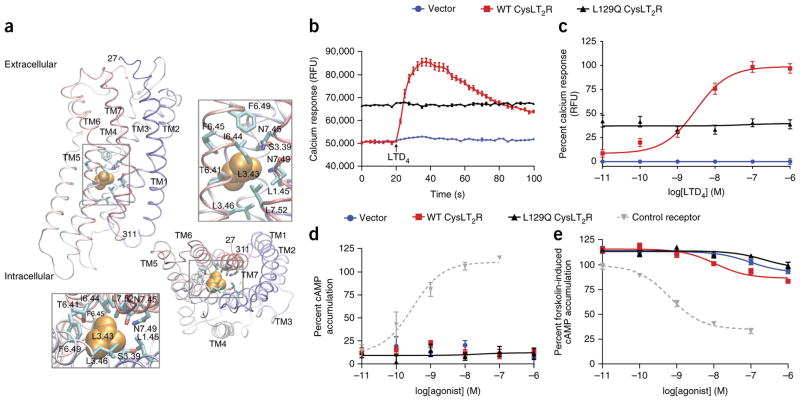
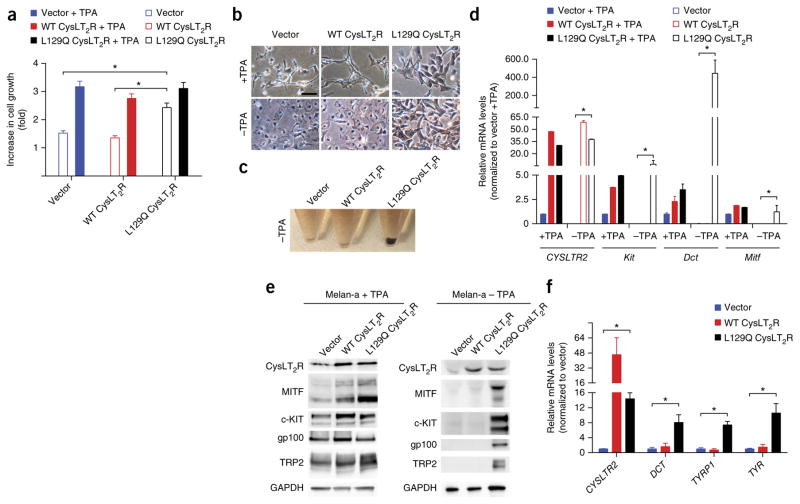
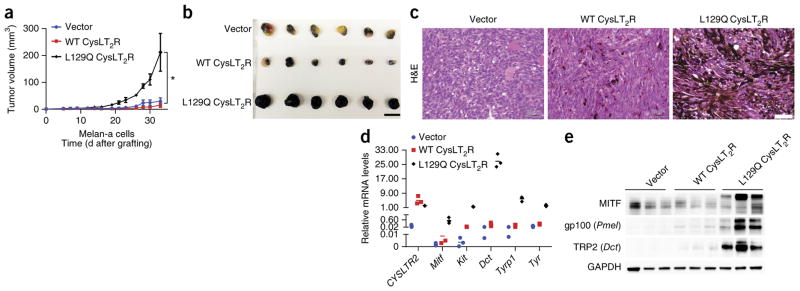
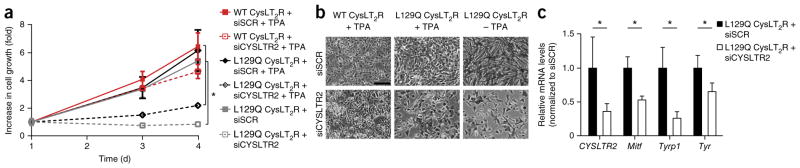
Uveal melanomas are molecularly distinct from cutaneous melanomas and lack mutations in BRAF, NRAS, KIT, and NF1. Instead, they are characterized by activating mutations in GNAQ and GNA11, two highly homologous α subunits of Gαq/11 heterotrimeric G proteins, and in PLCB4 (phospholipase C β4), the downstream effector of Gαq signaling. We analyzed genomics data from 136 uveal melanoma samples and found a recurrent mutation in CYSLTR2 (cysteinyl leukotriene receptor 2) encoding a p.Leu129Gln substitution in 4 of 9 samples that lacked mutations in GNAQ, GNA11, and PLCB4 but in 0 of 127 samples that harbored mutations in these genes. The Leu129Gln CysLT2R mutant protein constitutively activates endogenous Gαq and is unresponsive to stimulation by leukotriene. Expression of Leu129Gln CysLT2R in melanocytes enforces expression of a melanocyte-lineage signature, drives phorbol ester-independent growth in vitro, and promotes tumorigenesis in vivo. Our findings implicate CYSLTR2 as a uveal melanoma oncogene and highlight the critical role of Gαq signaling in uveal melanoma pathogenesis.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
