

Goblet cells of the conjunctiva: A review of recent findings
- PMID: 27091323
- PMCID: PMC4992623
- DOI: 10.1016/j.preteyeres.2016.04.005
Goblet cells of the conjunctiva: A review of recent findings
Abstract
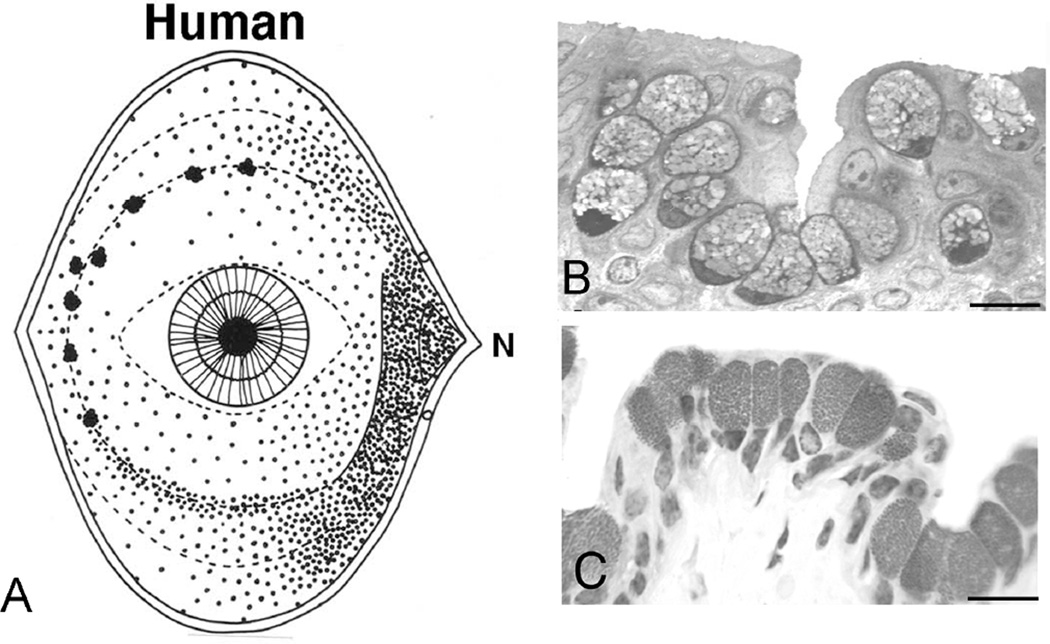
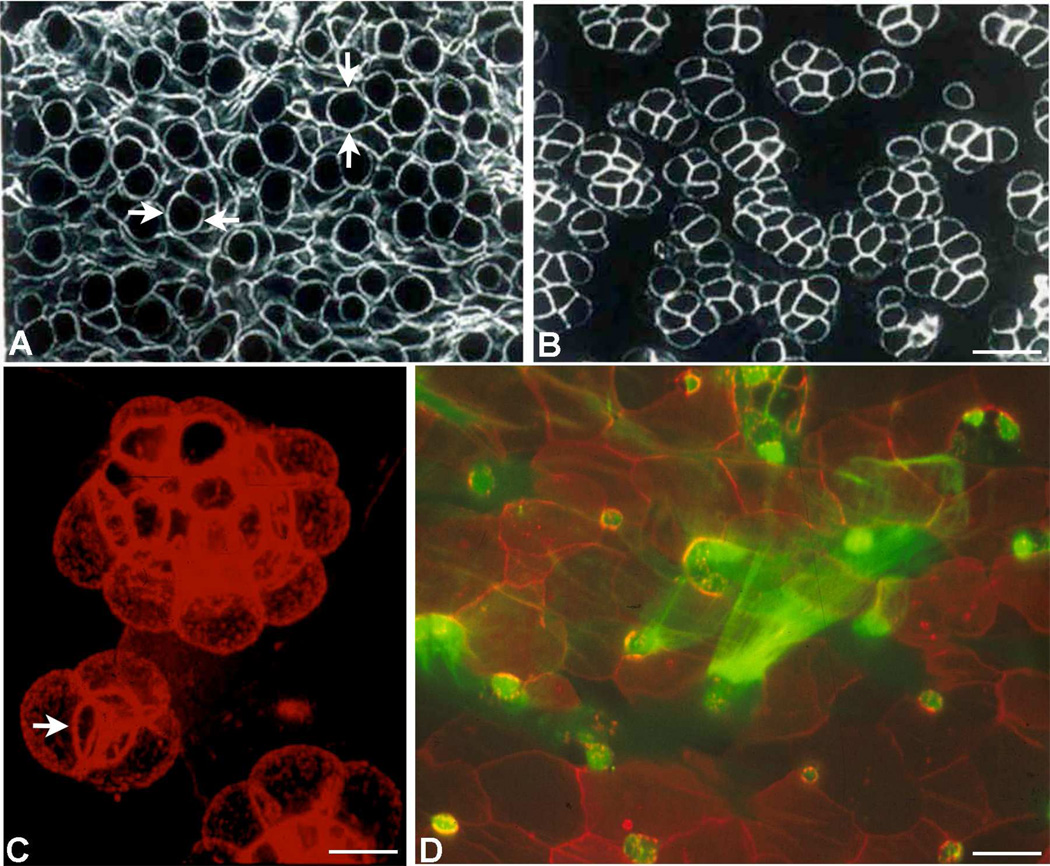
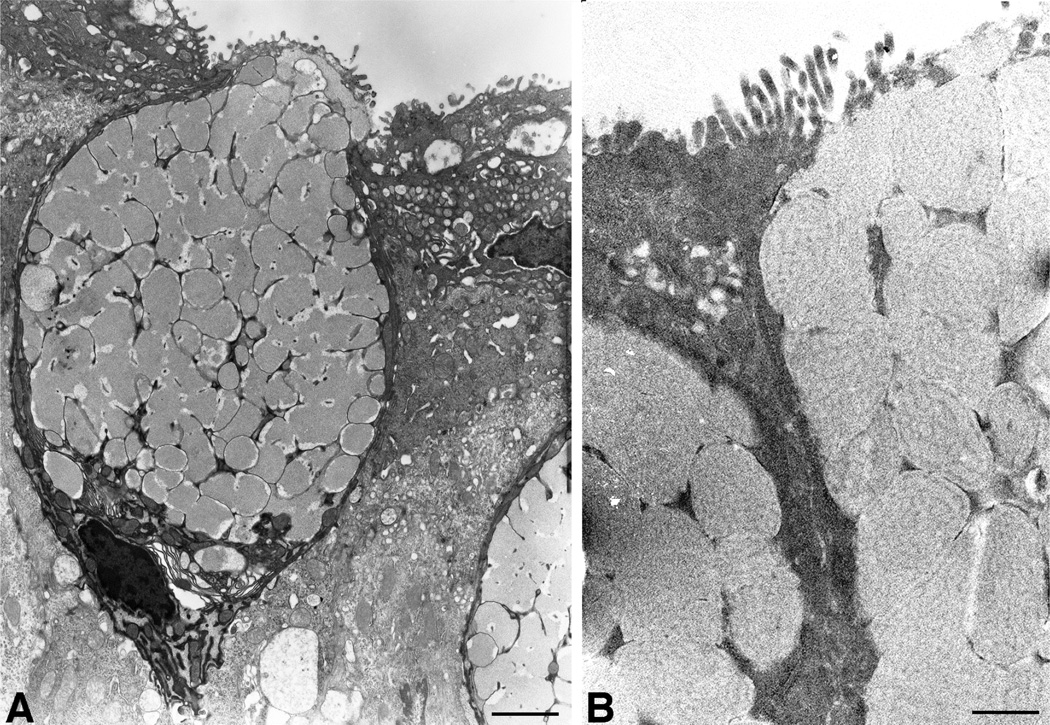
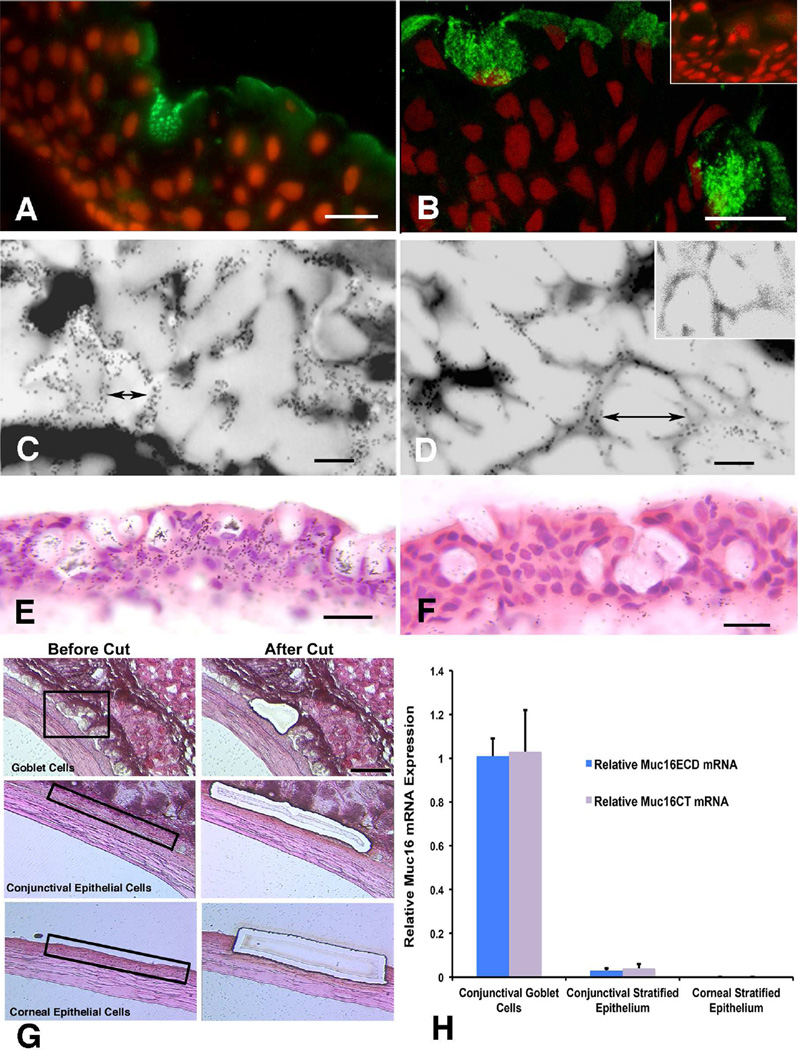
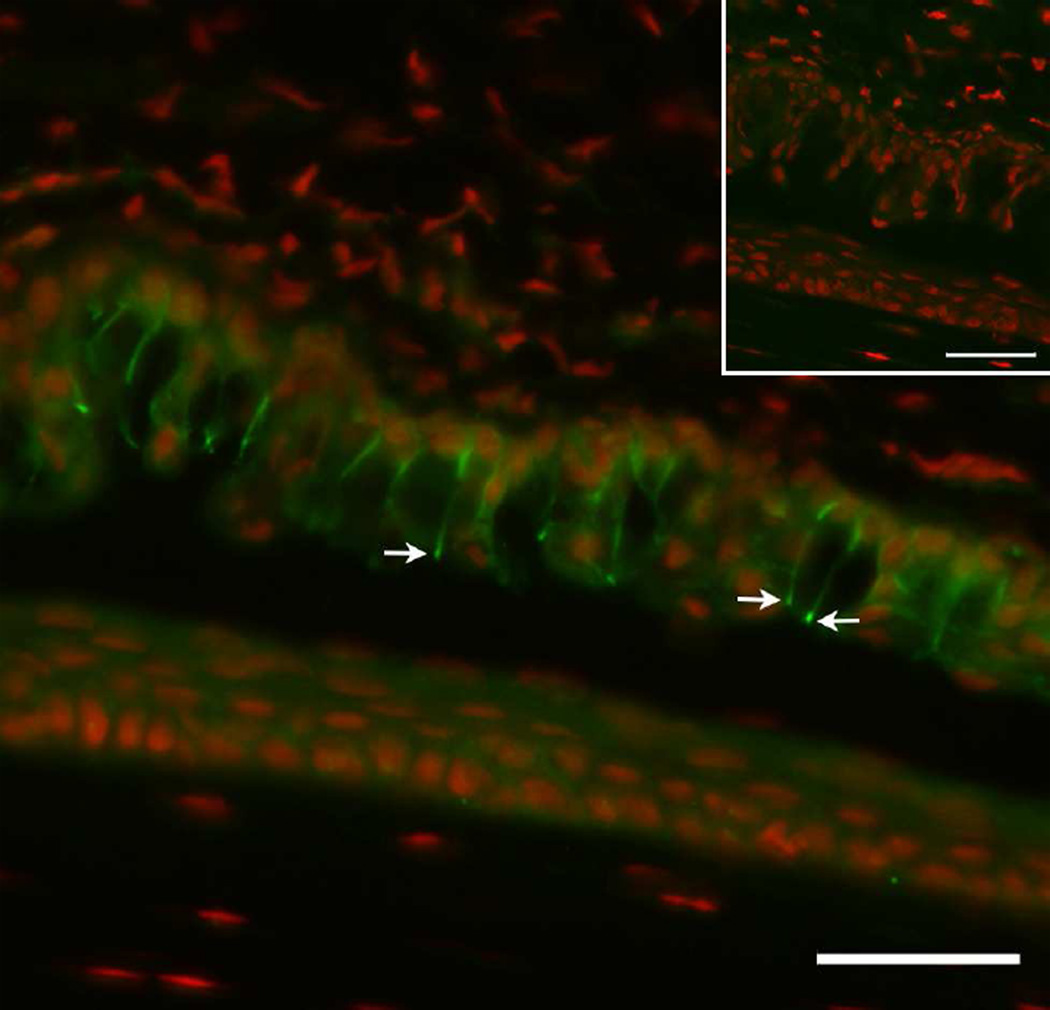
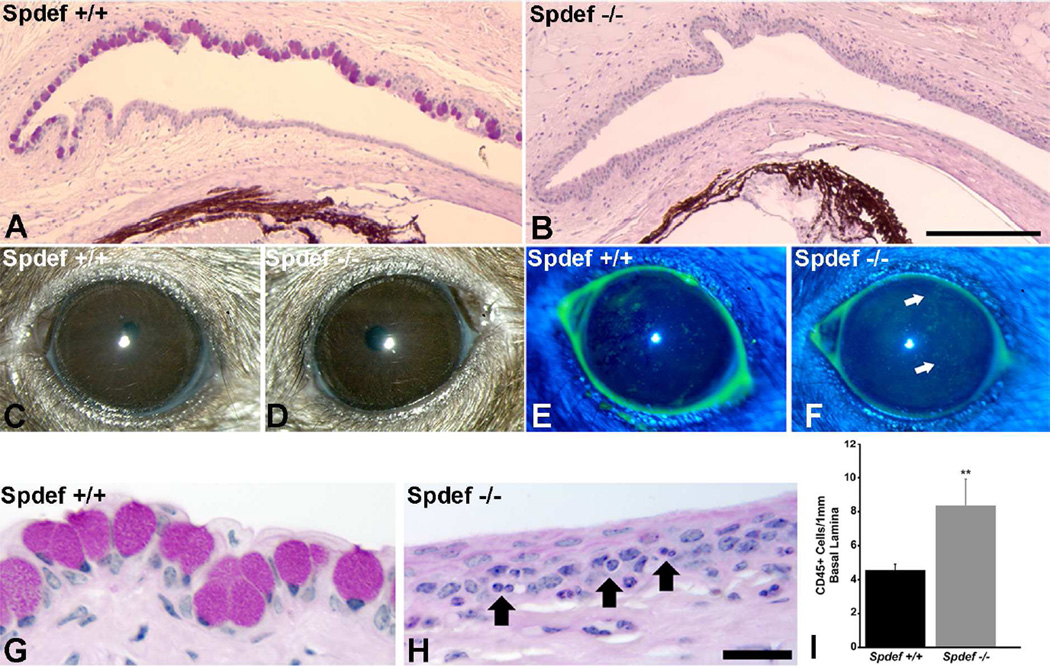
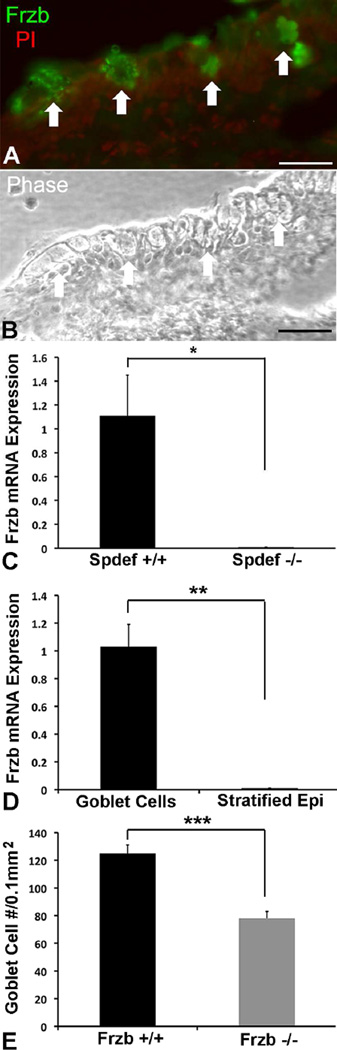
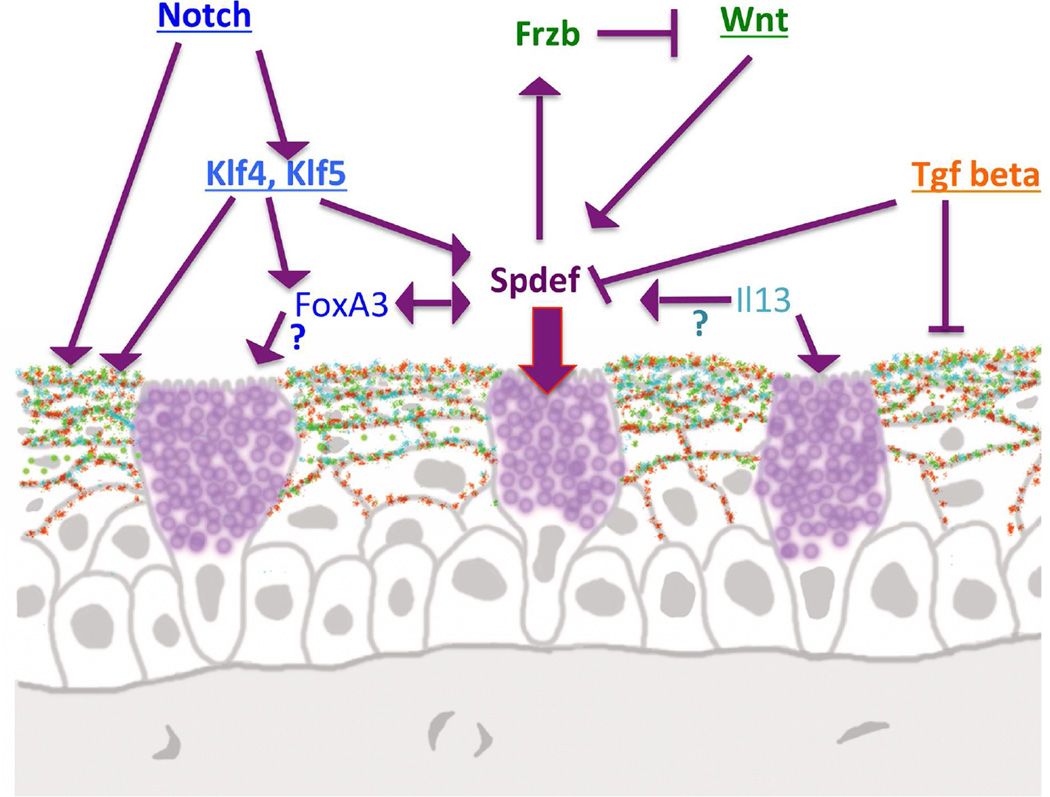
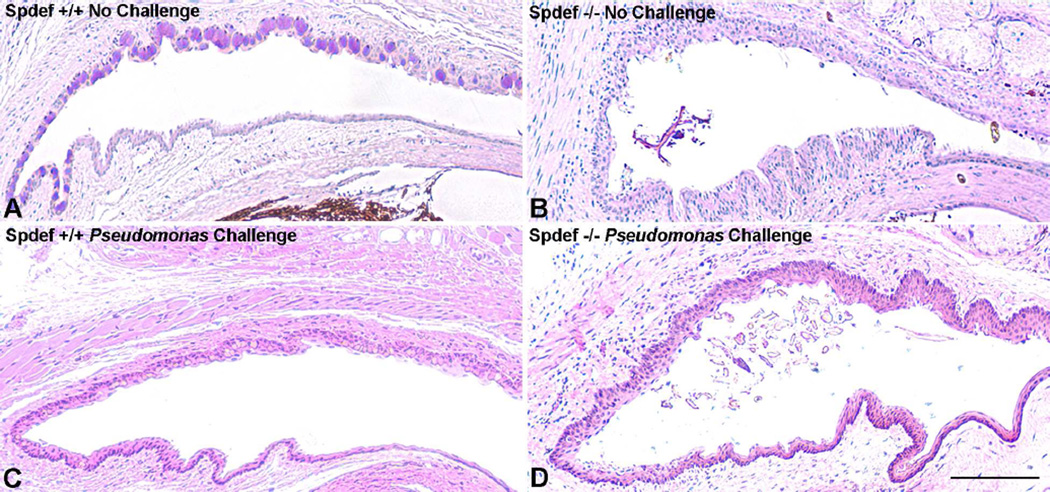
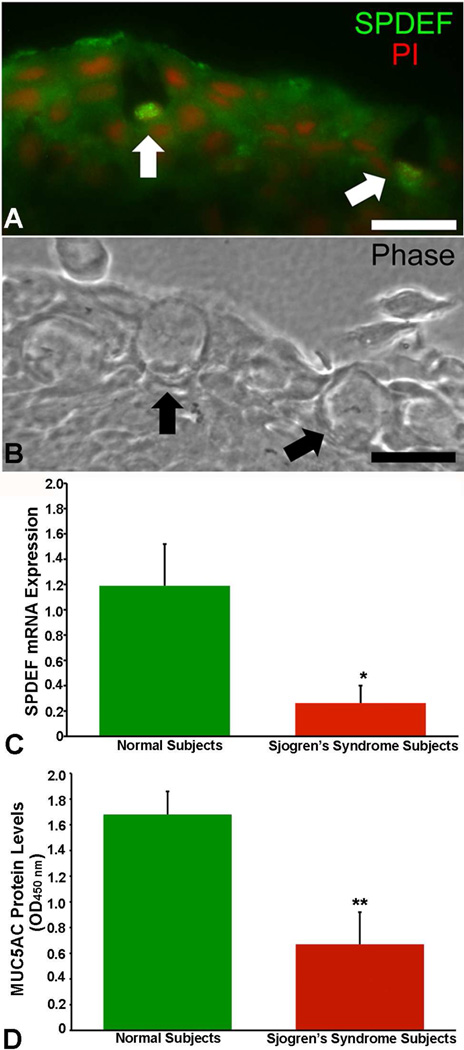
Goblet cells within the conjunctival epithelium are specialized cells that secrete mucins onto the surface of the eye. Recent research has demonstrated new characteristics of the cells, including factors influencing their differentiation, their gene products and their functions at the ocular surface. The following review summarizes the newly discovered aspects of the role of Spdef, a member of the Ets transcription factor family in conjunctival goblet cell differentiation, the newly discovered goblet cell products including claudin2, the Wnt inhibitor Frzb, and the transmembrane mucin Muc16. The current concepts of conjunctival goblet cell function, including debris removal and immune surveillance are reviewed, as are changes in the goblet cell population in ocular surface diseases. Major remaining questions regarding conjunctival cell biology are discussed.
Keywords: Conjunctival epithelium; Dry eye; Goblet cell; Goblet cell differentiation; Ocular surface; SPDEF.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures
References
-
- Argueso P, Balaram M, Spurr-Michaud S, Keutmann HT, Dana MR, Gipson IK. Decreased levels of the goblet cell mucin MUC5AC in tears of patients with Sjögren’s syndrome. Invest Ophthalmol Vis Sci. 2002;43:1004–1011. - PubMed
-
- Argueso P, Spurr-Michaud S, Russo CL, Tisdale A, Gipson IK. MUC16 mucin is expressed by the human ocular surface epithelia and carries the H185 carbohydrate epitope. Investigative ophthalmology & visual science. 2003a;44:2487–2495. http://dx.doi.org/10.1167/iovs.02-0862. - DOI - PubMed
-
- Argueso P, Tisdale A, Mandel U, Letko E, Foster CS, Gipson IK. The cell-layer- and cell-type-specific distribution of GalNAc-transferases in the ocular surface epithelia is altered during keratinization. Investigative ophthalmology & visual science. 2003b;44:86–92. http://dx.doi.org/10.1167/iovs.02-0181. - DOI - PubMed
-
- Barker N, van de Wetering M, Clevers H. The intestinal stem cell. Genes & development. 2008;22:1856–1864. http://dx.doi.org/10.1101/gad.1674008. - DOI - PMC - PubMed
-
- Bell SM, Zhang L, Xu Y, Besnard V, Wert SE, Shroyer N, Whitsett JA. Kruppel-like factor 5 controls villus formation and initiation of cytodifferentiation in the embryonic intestinal epithelium. Developmental biology. 2013;375:128–139. http://dx.doi.org/10.1016/j.ydbio.2012.12.010. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
