

TREM2-mediated early microglial response limits diffusion and toxicity of amyloid plaques
- PMID: 27091843
- PMCID: PMC4854736
- DOI: 10.1084/jem.20151948
TREM2-mediated early microglial response limits diffusion and toxicity of amyloid plaques
Abstract
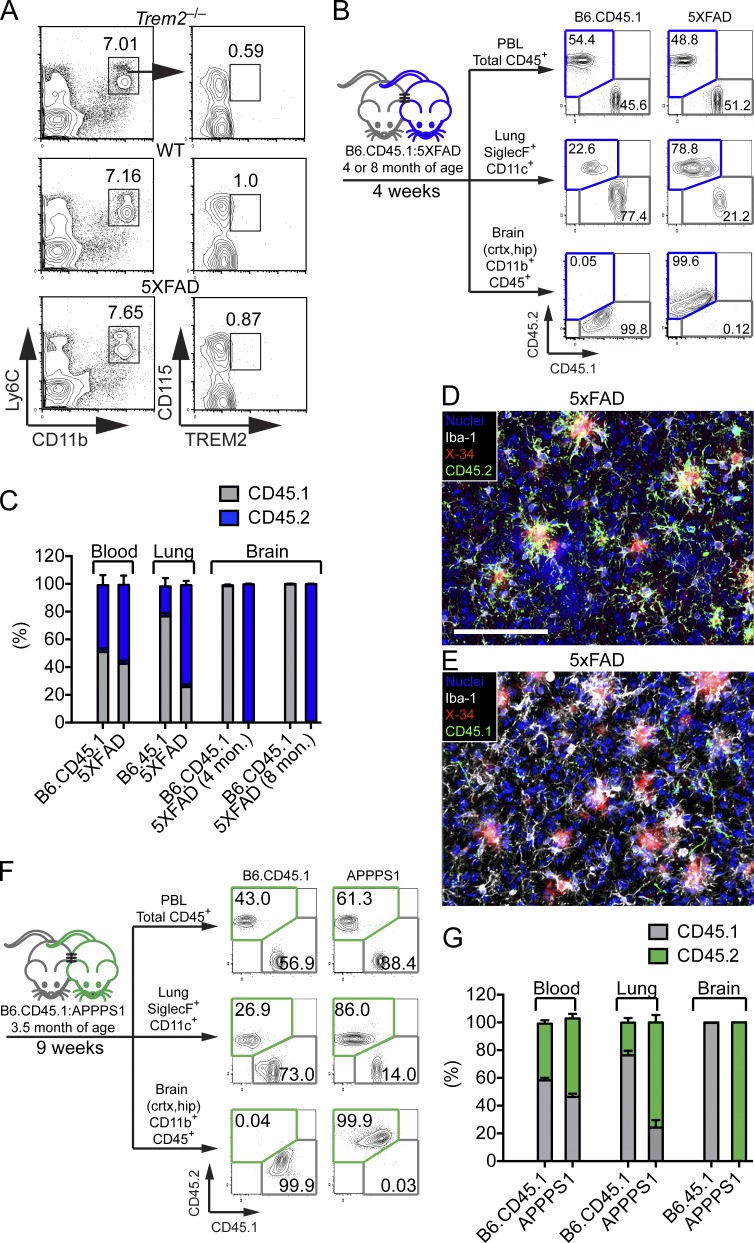
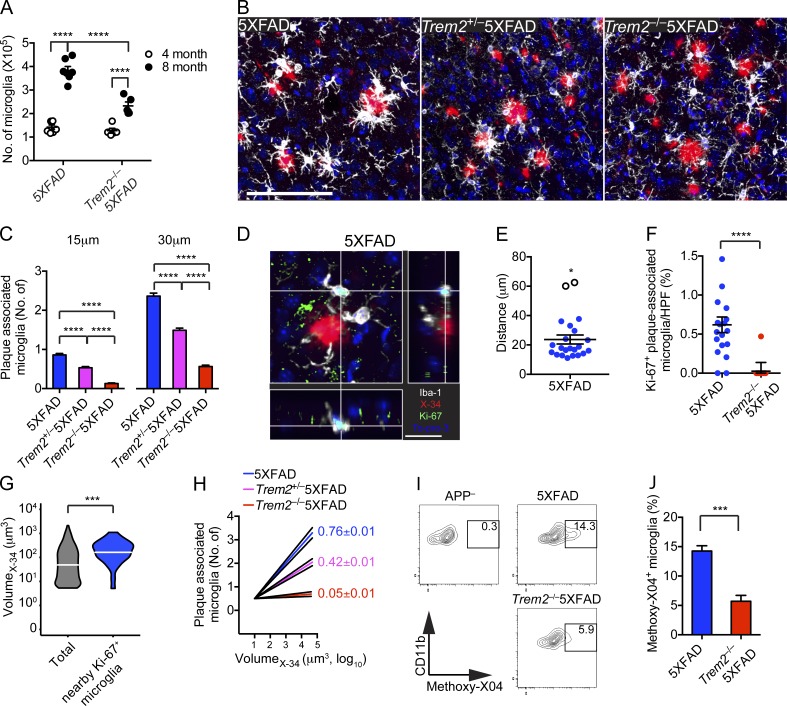
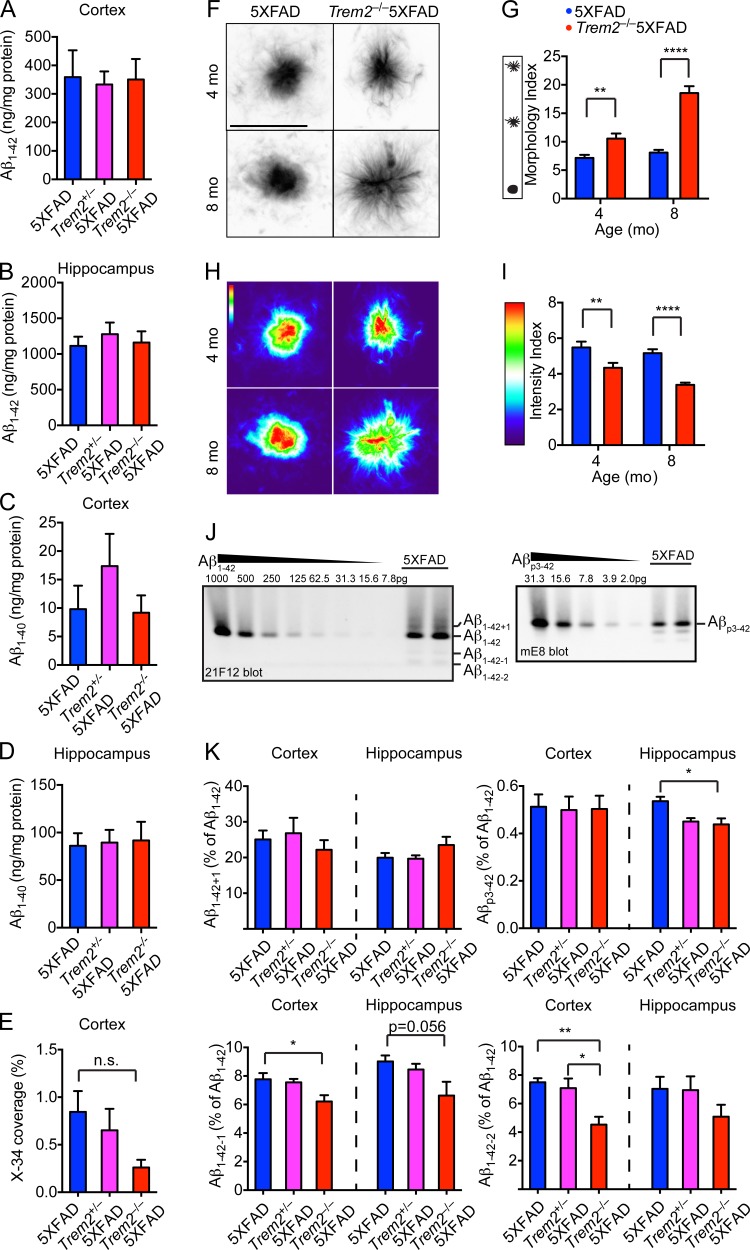
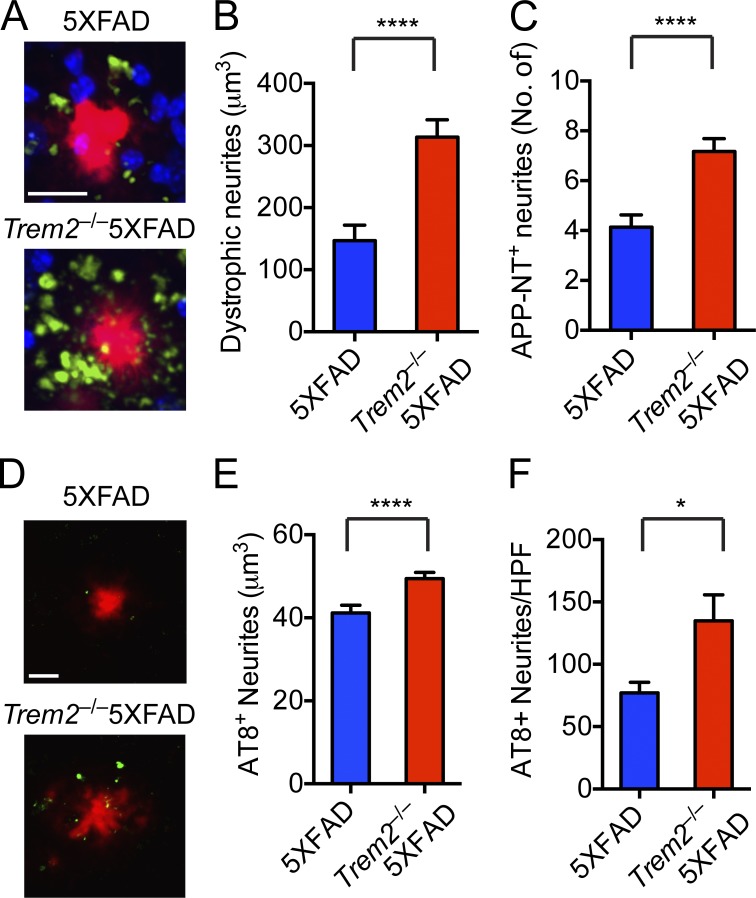
Triggering receptor expressed on myeloid cells 2 (TREM2) is a microglial receptor that recognizes changes in the lipid microenvironment, which may occur during amyloid β (Aβ) accumulation and neuronal degeneration in Alzheimer's disease (AD). Rare TREM2 variants that affect TREM2 function lead to an increased risk of developing AD. In murine models of AD, TREM2 deficiency prevents microglial clustering around Aβ deposits. However, the origin of myeloid cells surrounding amyloid and the impact of TREM2 on Aβ accumulation are a matter of debate. Using parabiosis, we found that amyloid-associated myeloid cells derive from brain-resident microglia rather than from recruitment of peripheral blood monocytes. To determine the impact of TREM2 deficiency on Aβ accumulation, we examined Aβ plaques in the 5XFAD model of AD at the onset of Aβ-related pathology. At this early time point, Aβ accumulation was similar in TREM2-deficient and -sufficient 5XFAD mice. However, in the absence of TREM2, Aβ plaques were not fully enclosed by microglia; they were more diffuse, less dense, and were associated with significantly greater neuritic damage. Thus, TREM2 protects from AD by enabling microglia to surround and alter Aβ plaque structure, thereby limiting neuritic damage.
© 2016 Wang et al.
Figures
References
-
- Brendza R.P., O’Brien C., Simmons K., McKeel D.W., Bales K.R., Paul S.M., Olney J.W., Sanes J.R., and Holtzman D.M.. 2003. PDAPP; YFP double transgenic mice: a tool to study amyloid-beta associated changes in axonal, dendritic, and synaptic structures. J. Comp. Neurol. 456:375–383. 10.1002/cne.10536 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
