

Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize
- PMID: 27092004
- PMCID: PMC4983849
- DOI: 10.1073/pnas.1601352113
Gene duplication confers enhanced expression of 27-kDa γ-zein for endosperm modification in quality protein maize
Abstract
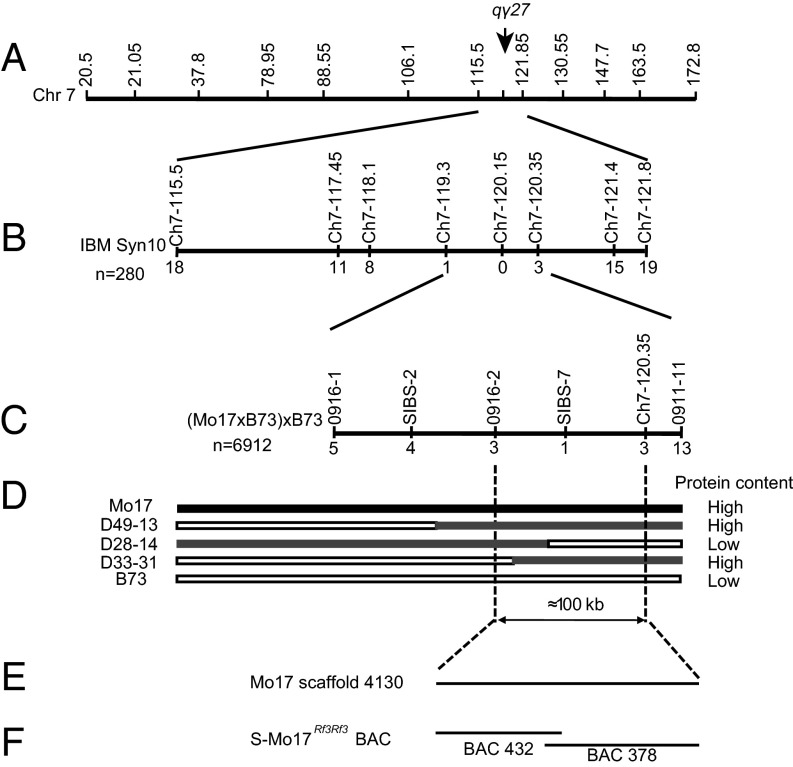
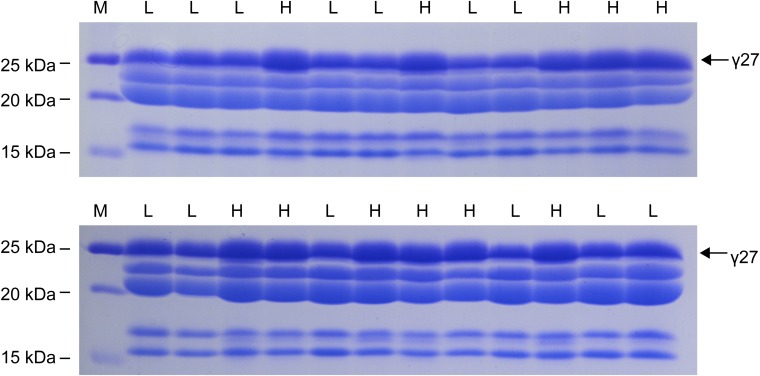
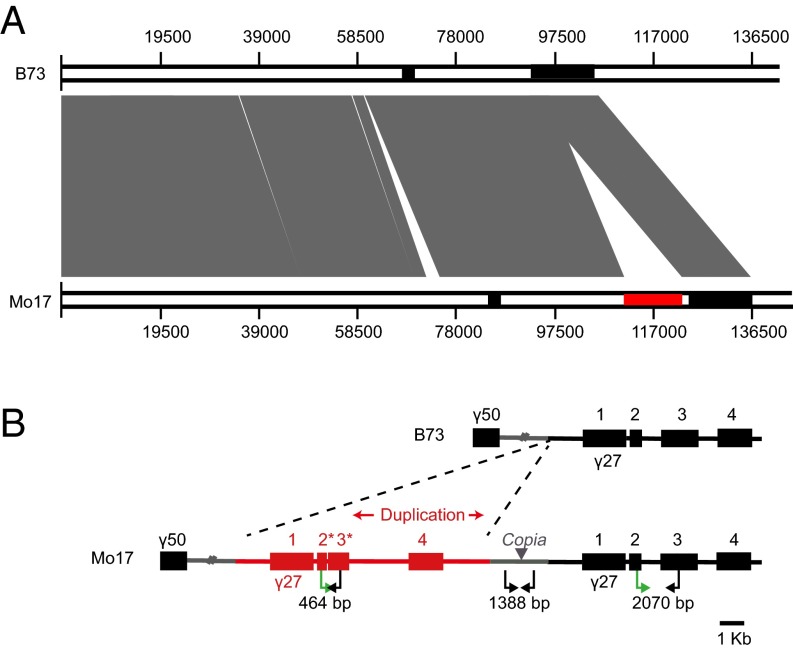
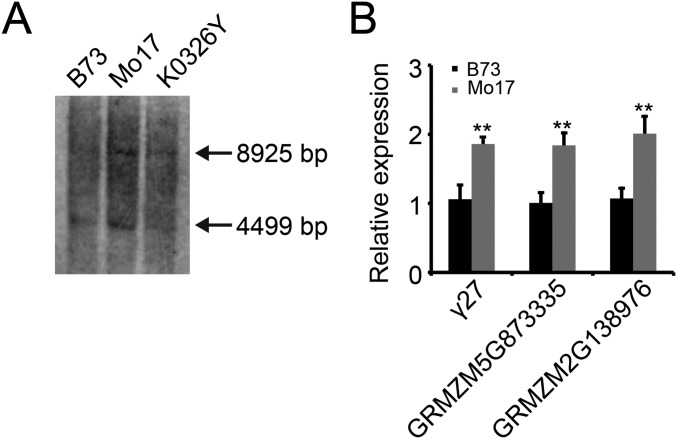
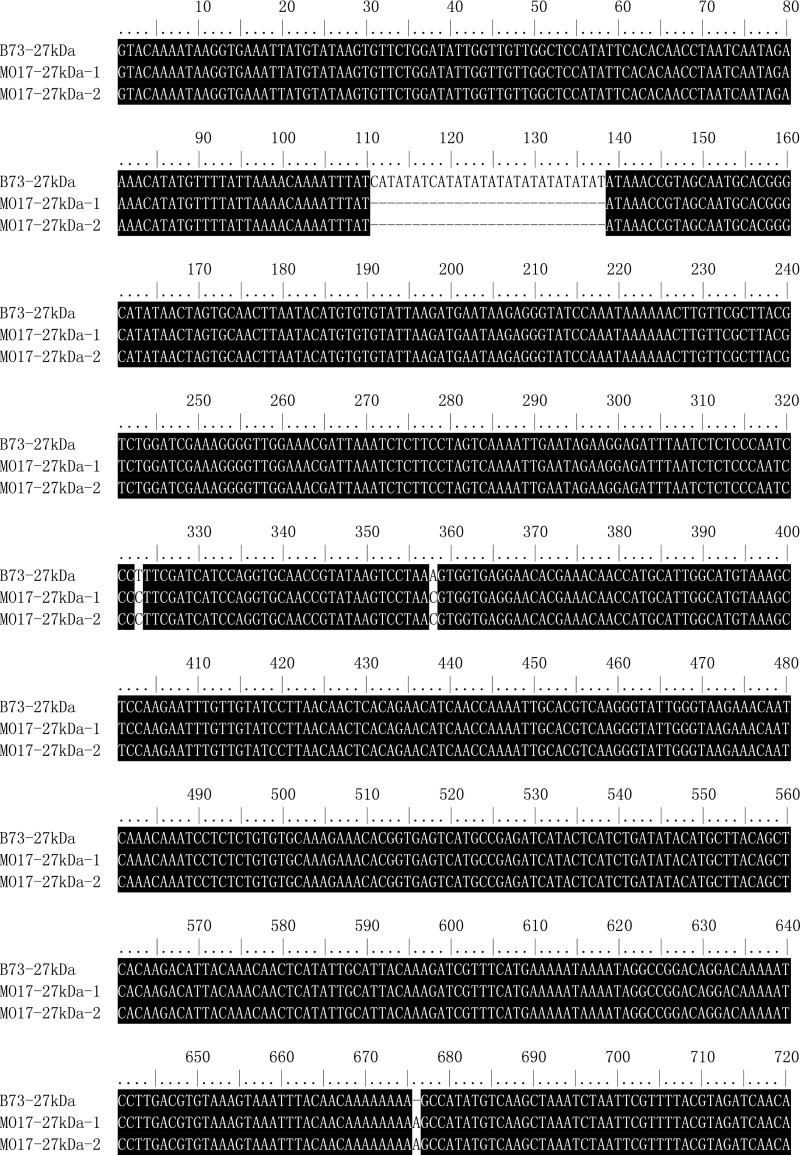
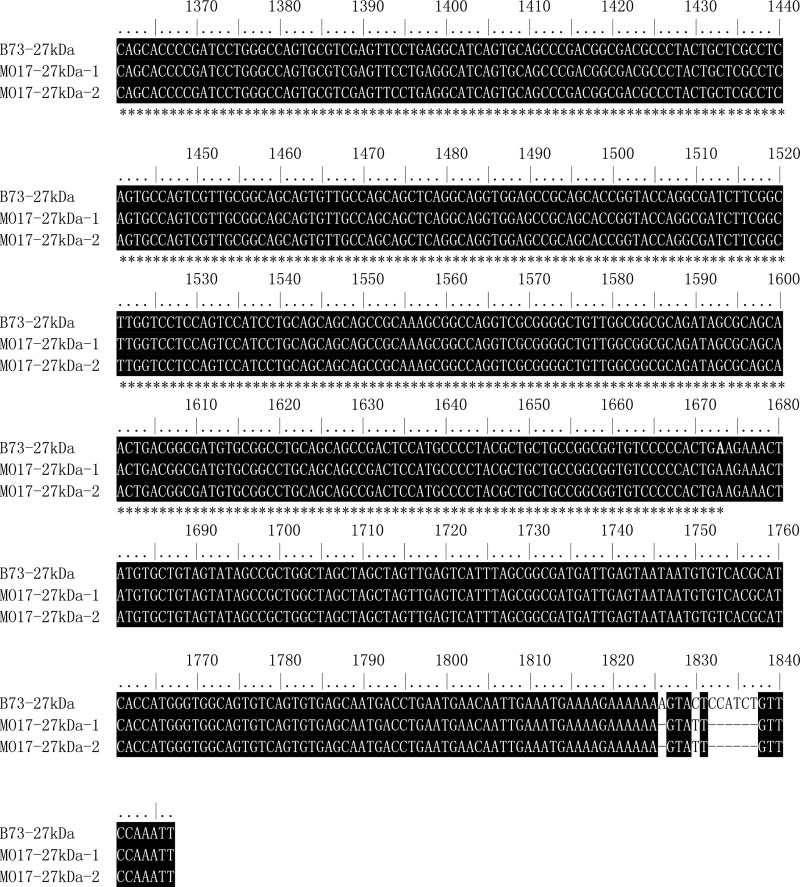
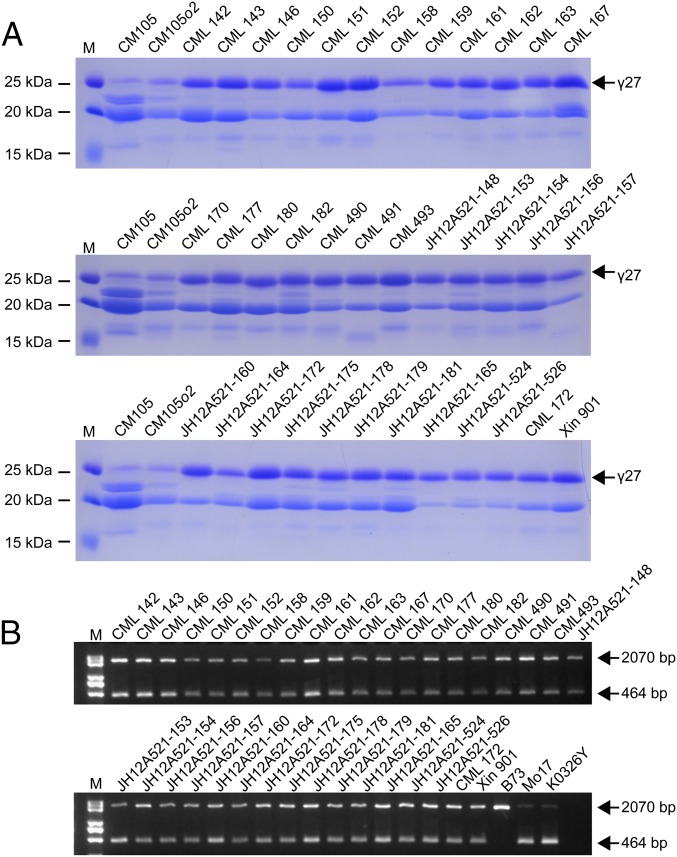
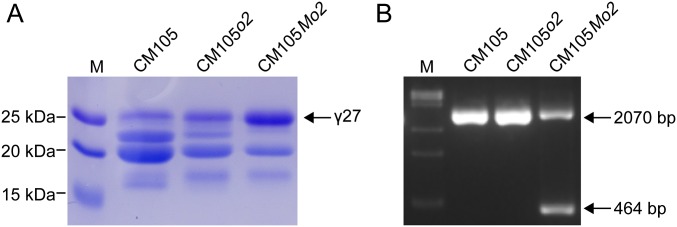
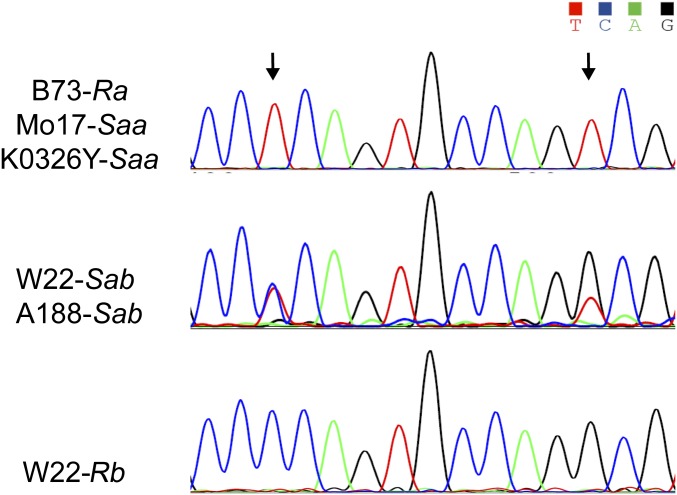
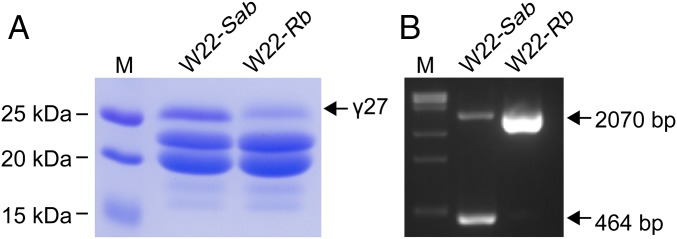
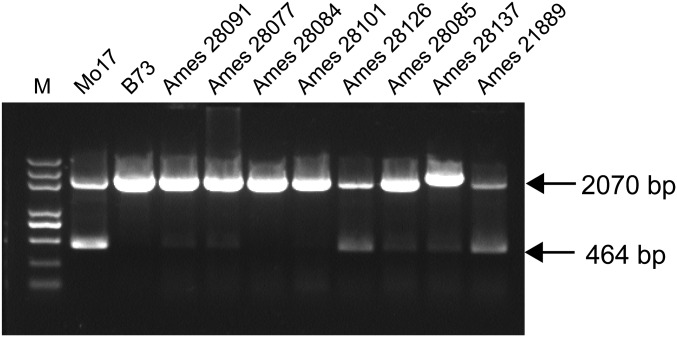
The maize opaque2 (o2) mutant has a high nutritional value but it develops a chalky endosperm that limits its practical use. Genetic selection for o2 modifiers can convert the normally chalky endosperm of the mutant into a hard, vitreous phenotype, yielding what is known as quality protein maize (QPM). Previous studies have shown that enhanced expression of 27-kDa γ-zein in QPM is essential for endosperm modification. Taking advantage of genome-wide association study analysis of a natural population, linkage mapping analysis of a recombinant inbred line population, and map-based cloning, we identified a quantitative trait locus (qγ27) affecting expression of 27-kDa γ-zein. qγ27 was mapped to the same region as the major o2 modifier (o2 modifier1) on chromosome 7 near the 27-kDa γ-zein locus. qγ27 resulted from a 15.26-kb duplication at the 27-kDa γ-zein locus, which increases the level of gene expression. This duplication occurred before maize domestication; however, the gene structure of qγ27 appears to be unstable and the DNA rearrangement frequently occurs at this locus. Because enhanced expression of 27-kDa γ-zein is critical for endosperm modification in QPM, qγ27 is expected to be under artificial selection. This discovery provides a useful molecular marker that can be used to accelerate QPM breeding.
Keywords: QPM; artificial selection; endosperm; gene duplication; o2 modifiers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
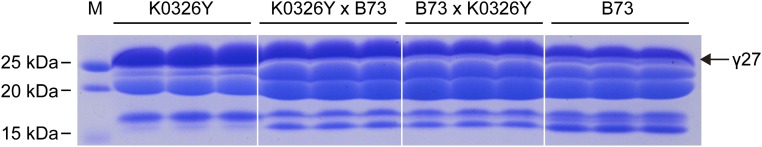
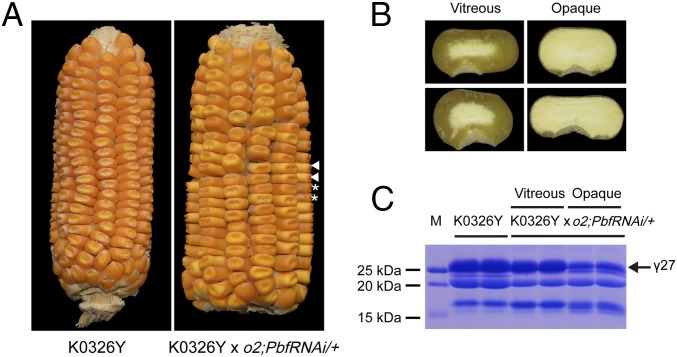
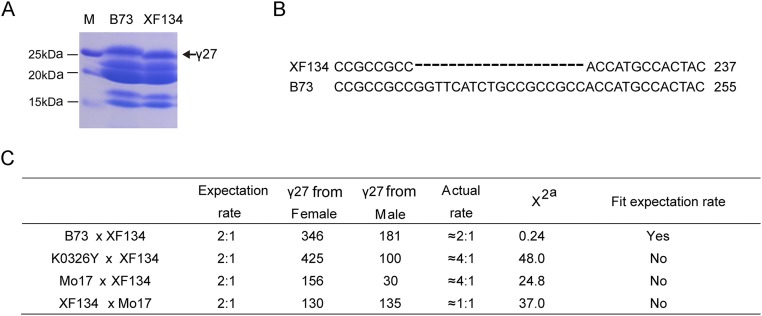
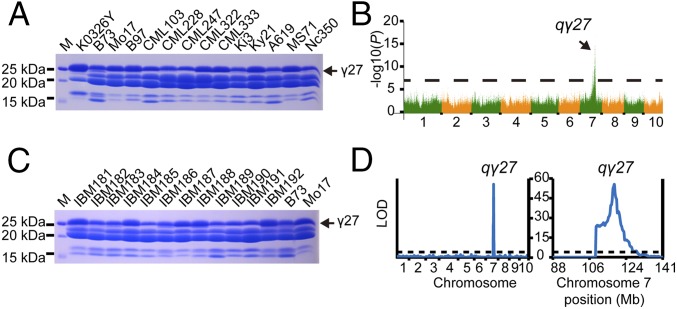

Similar articles
-
Deletion mutagenesis identifies a haploinsufficient role for γ-zein in opaque2 endosperm modification.Plant Physiol. 2014 Jan;164(1):119-30. doi: 10.1104/pp.113.230961. Epub 2013 Nov 8. Plant Physiol. 2014. PMID: 24214534 Free PMC article.
-
opaque2 modifiers act post-transcriptionally and in a polar manner on gamma-zein gene expression in maize endosperm.Plant Cell. 1993 Nov;5(11):1599-609. doi: 10.1105/tpc.5.11.1599. Plant Cell. 1993. PMID: 8312742 Free PMC article.
-
Characterization of opaque2 modifier QTLs and candidate genes in recombinant inbred lines derived from the K0326Y quality protein maize inbred.Theor Appl Genet. 2011 Mar;122(4):783-94. doi: 10.1007/s00122-010-1486-3. Epub 2010 Nov 13. Theor Appl Genet. 2011. PMID: 21076810
-
Regulation of alpha-zein gene expression during maize endosperm development.Results Probl Cell Differ. 1994;20:209-33. doi: 10.1007/978-3-540-48037-2_10. Results Probl Cell Differ. 1994. PMID: 8036317 Review. No abstract available.
-
Molecular mechanisms regulating the synthesis of storage proteins in maize endosperm.Crit Rev Food Sci Nutr. 1982;16(2):199-215. doi: 10.1080/10408398209527334. Crit Rev Food Sci Nutr. 1982. PMID: 7039978 Review. No abstract available.
Cited by
-
Nitrogen-dependent binding of the transcription factor PBF1 contributes to the balance of protein and carbohydrate storage in maize endosperm.Plant Cell. 2023 Jan 2;35(1):409-434. doi: 10.1093/plcell/koac302. Plant Cell. 2023. PMID: 36222567 Free PMC article.
-
Variance of Zein Protein and Starch Granule Morphology between Corn and Steam Flaked Products Determined Starch Ruminal Degradability Through Altering Starch Hydrolyzing Bacteria Attachment.Animals (Basel). 2019 Aug 29;9(9):626. doi: 10.3390/ani9090626. Animals (Basel). 2019. PMID: 31470611 Free PMC article.
-
Maize opaque mutants are no longer so opaque.Plant Reprod. 2018 Sep;31(3):319-326. doi: 10.1007/s00497-018-0344-3. Epub 2018 Jul 5. Plant Reprod. 2018. PMID: 29978299 Free PMC article. Review.
-
Quantitative Trait Locus Analysis for Deep-Sowing Germination Ability in the Maize IBM Syn10 DH Population.Front Plant Sci. 2017 May 22;8:813. doi: 10.3389/fpls.2017.00813. eCollection 2017. Front Plant Sci. 2017. PMID: 28588594 Free PMC article.
-
A genome-wide association study identifies a transporter for zinc uploading to maize kernels.EMBO Rep. 2023 Jan 9;24(1):e55542. doi: 10.15252/embr.202255542. Epub 2022 Nov 17. EMBO Rep. 2023. PMID: 36394374 Free PMC article.
References
-
- Wheeler T, von Braun J. Climate change impacts on global food security. Science. 2013;341(6145):508–513. - PubMed
-
- Wu Y, Yuan L, Guo X, Holding DR, Messing J. Mutation in the seed storage protein kafirin creates a high-value food trait in sorghum. Nat Commun. 2013;4:2217. - PubMed
-
- Gilding EK, et al. Allelic variation at a single gene increases food value in a drought-tolerant staple cereal. Nat Commun. 2013;4:1483. - PubMed
-
- Osborne TB, Mendel LB; With the cooperation of Edna LF and Alfred JW (1914) Nutritive properties of proteins of the maize kernel. J Biol Chem 18(1):1–16.
-
- Vasal SK, Villegas E, Bjarnason M, Gelaw B, Goertz P. Genetic modifiers and breeding strategies in developing hard endosperm opaque-2 materials. In: Pollmer WG, Phillips RH, editors. Improvement of Quality Traits of Maize for Grain and Silage Use. Martinus Nijhoff; London: 1980. pp. 37–73.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
