

Review
doi: 10.1038/nrm.2016.36.
Epub 2016 Apr 20.
Hitchhiking on the heptahelical highway: structure and function of 7TM receptor complexes
Affiliations
- PMID: 27093944
- PMCID: PMC5503103
- DOI: 10.1038/nrm.2016.36
Item in Clipboard
Review
Hitchhiking on the heptahelical highway: structure and function of 7TM receptor complexes
Nat Rev Mol Cell Biol.
2016 Jul.
Abstract
A revolution in the analysis of seven transmembrane domain (7TM) receptors has provided detailed information about how these physiologically important signalling proteins interact with extracellular cues. However, it has proved much more challenging to understand how 7TM receptors convey information to their principal intracellular targets: heterotrimeric G proteins, G protein-coupled receptor kinases and arrestins. Recent structures now suggest a common mechanism that enables these structurally diverse cytoplasmic proteins to 'hitch a ride' on hundreds of different activated 7TM receptors in order to instigate physiological change.
Conflict of interest statement
The author declares no competing interests.
Figures
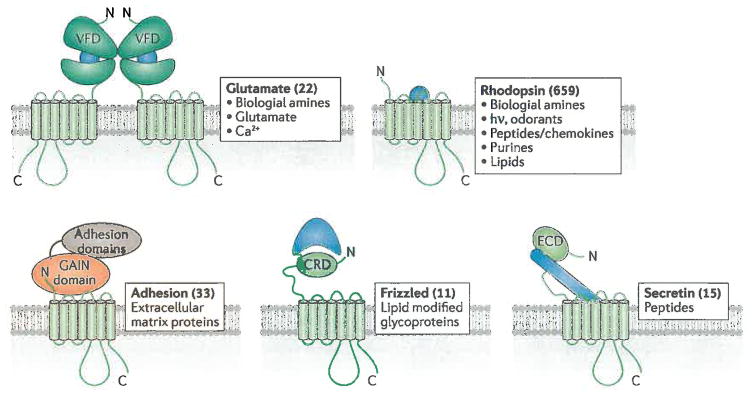
7TM receptor families and their ligands. 7TM receptor families all possess a characteristic arrangement of 7 transmembrane spans, but often diverse extracellular domains. Numbers in parentheses correspond to the number of receptors identified in each family in humans and some representative activating lignads are listed for each group. Odorant receptors (388 in number) are included in the rhodopsin family. Not shown are the taste2 receptors (25 in number), whose transmembrane domains most closely resemble those of the frizzled family, but lack analogous extracellular domains. Glutamate family receptors dimerize through their VFD domains. The GAIN domain of adhesion receptors has autoproteolytic activity that cleaves its extracellular domain such that the mature protein exists as two non-covalently associated subunits. Blue shapes represent the mode by which agonists interact with the extracellular regions in each receptor family. The frizzled ligand is covalently modified by a palmitoyl group (jagged line), which forms part of the interface with the CRD domain. VFD, Venus fly-trap domain; CRD, cysteine-rich domain; ECD, extracellular domain; GAIN, GPCR autoproteolysis inducing.
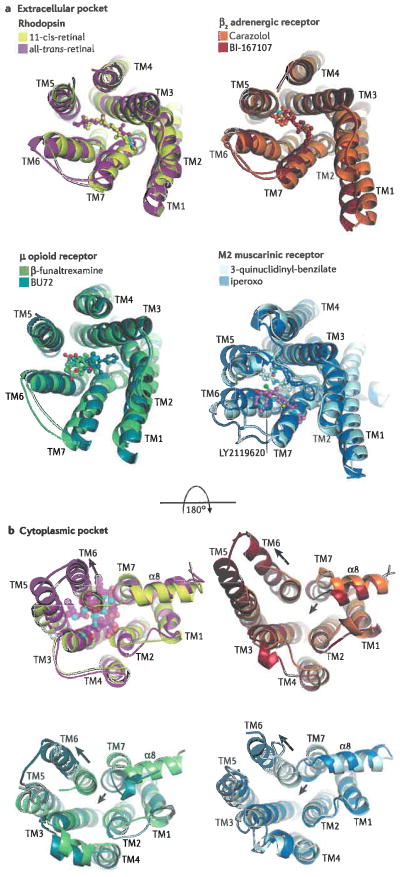
Extracellular (a) and cytoplasmic (b) pockets of four different 7TM receptors: rhodopsin (PDB entries 1F88, yellow, and 3PQR, magenta), the β2AR (PDB entries 2RH1, orange, and 3SN6, brick), the μ-opioid receptor (PDB entries 4DKL, chartreuse, and 5C1M, teal), and the M2 muscarinic receptor (PDB entries 3UON, pale cyan, and 4MQT, blue). In each overlay, the darker colour corresponds to an active, agonist-bound form of the receptor, whereas the lighter colour corresponds to an inactive form. Panel a illustrates the extracellular pockets bound to various ligands (ball and stick models, with carbons coloured the same as their receptor). N-termini and the second extracellular loop (the loop connecting TM4 and TM5) were removed when necessary for clarity. Photoactivation or the binding of ligands leads to relatively subtle and inconsistent changes in the helices and loops that form the extracellular pocket (rhodopsin expands, β2AR is relatively unchanged, the μ-opioid and M2 receptors contract). LY2119620 (ball and stick model with pink carbons; see muscarinic receptor) is a positive allosteric modulator that binds to the ‘vestibule’ of the M2 muscarinic receptor, formed by the three extracellular loops of the receptor, and thereby increases the effective affinity of the agonist iperoxo. Receptor activation promotes more dramatic yet consistent changes in the cytoplasmic pockets (illustrated in panel b), where the end of TM6 swings away from the core of the transmembrane domain (long arrows), and TM7 and the α8 helix tend to push inward (short arrows) except in the case of rhodopsin. This transition creates a binding site for cytoplasmic proteins, as represented by the transducin peptide bound to opsin (transparent spheres in the cytoplasmic pocket of rhodopsin).
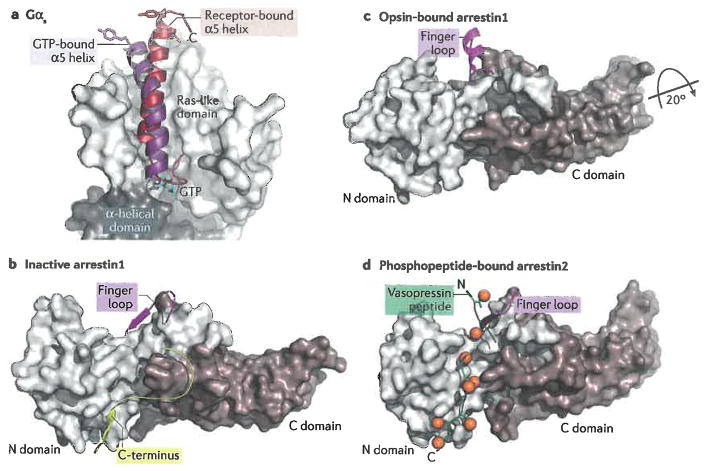
Interactions of heterotrimeric G proteins and arrestins with 7TM receptors. a) Schematic summarizing how Gα subunits bind to activated receptors. The most prominent interaction is that of its C-terminus (α5) with the cytoplasmic pocket of the receptor. This results in lowering affinity for bound nucleotides and allows the α helical domain to swing away from the Ras-like domain, facilitating nucleotide exchange . Arrows represent the β-strands that form the core sheet of the Ras-like domain. The β1 strand, which precedes the P loop that binds phosphate moieties of guanine nucleotides (circles marked with “P”), has also been implicated in become more solvent accessible in the nucleotide free state. The termini of Gα are marked N and C. b) The C-terminal α5 helix of Ras-like domain of Gαs features a markedly different position in the receptor bound conformation (firebrick, PDB entry 3SN6) as compared to its conformation in its GTP-bound state (purple, PDB entry 1AZT). The side chains of two hydrophobic residues (Leu388 and Tyr391) that make extensive interactions with TM3, TM5 and TM6 of the receptor (in this case β2AR) are shown as sticks to emphasize the change in the position of α5 helix. c) Schematic of the interactions of arrestin with an activated receptor. Most contacts are formed by the N domain, most prominently by the finger loop which projects into the cytoplasmic pocket. The N domain also forms most of the interactions with phosphorylated residues from the receptor. Together these interactions help to rearrange the polar core between the N and C domains and release the C-terminus of arrestin from the N domain. d) Inactive structure of arrestin1 (PDB entry 1CF1). Its C-terminus (yellow) binds to the N-domain, thereby blocking interactions with MAP kinases and with the endocytotic machinery. e) When arrestin binds to an activated receptor, the C-domain twists ~20° relative to the N-domain and the conformation of the finger loop (magenta)is altered so that it can directly interact with the transmembrane domains of 7TM receptors (PDB entry 4ZWJ ). f) Arrestins also bind to the phosphorylated C-terminal tails of 7TM receptors, as exemplified here by the arrestin2-vasopressin phosphopeptide complex (PDB entry 4JQI ). Phosphates are depicted as orange spheres. Note that the C-terminal end of the vasopressin peptide would displace the C-terminus of arrestin shown in panel b indicating that this interaction can activate the signalling function of arrestins by unblocking the sites on the N domain responsible for interactions with other cellular components. It remains to be determined which of the “activated” finger loop models shown, if any, are physiologically relevant. The conformation observed in 4JQI could, however, reflect how arrestin2 binds to a phosphorylated C-terminal tail when it is not also engaged with the transmembrane domain of a 7TM receptor.
References
-
- Fredriksson R, Lagerstrom MC, Lundin LG, Schioth HB. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol Pharmacol. 2003;63:1256–72. - PubMed
-
- Bjarnadottir TK, et al. Comprehensive repertoire and phylogenetic analysis of the G protein-coupled receptors in human and mouse. Genomics. 2006;88:263–73. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
