

Nuclear factor erythroid-derived 2-like 2 (NFE2L2, Nrf2) mediates exercise-induced mitochondrial biogenesis and the anti-oxidant response in mice
- PMID: 27094017
- PMCID: PMC5023720
- DOI: 10.1113/JP271957
Nuclear factor erythroid-derived 2-like 2 (NFE2L2, Nrf2) mediates exercise-induced mitochondrial biogenesis and the anti-oxidant response in mice
Abstract
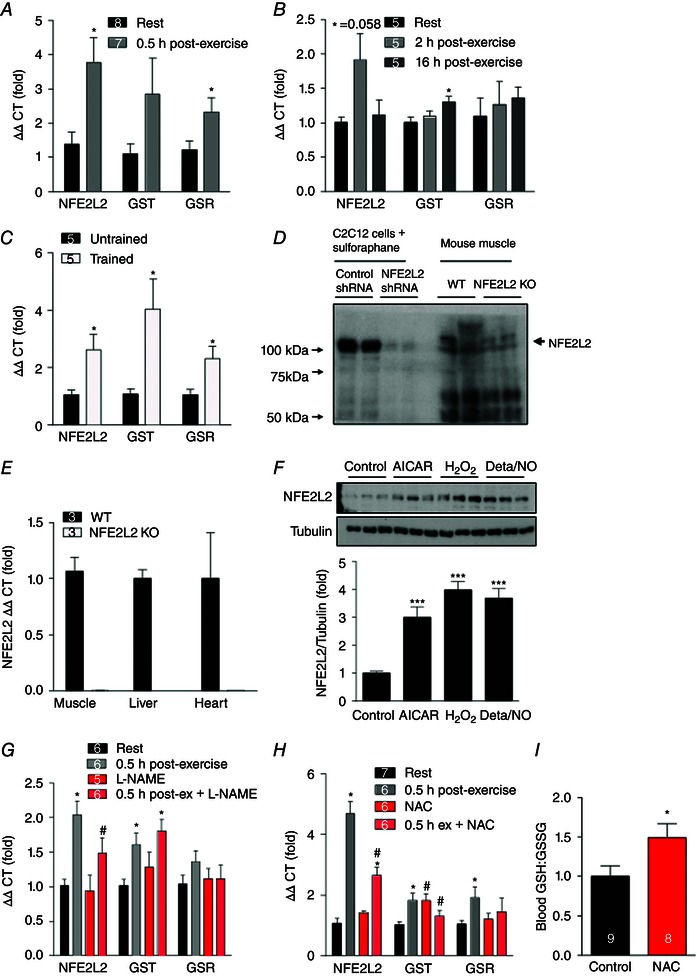
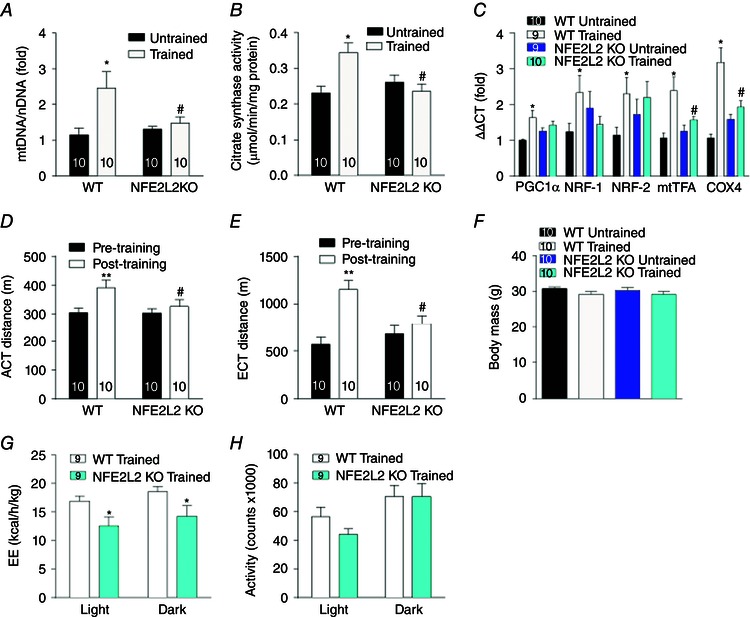
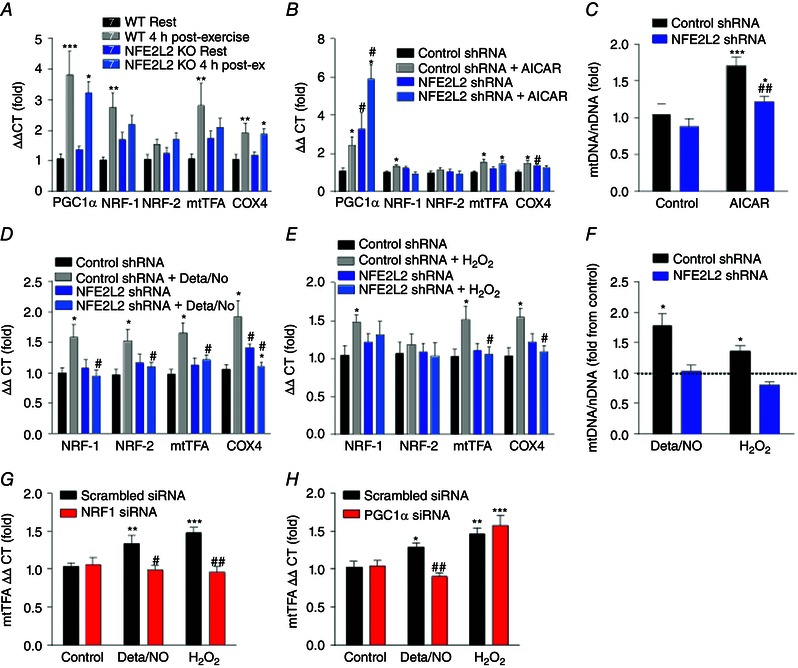
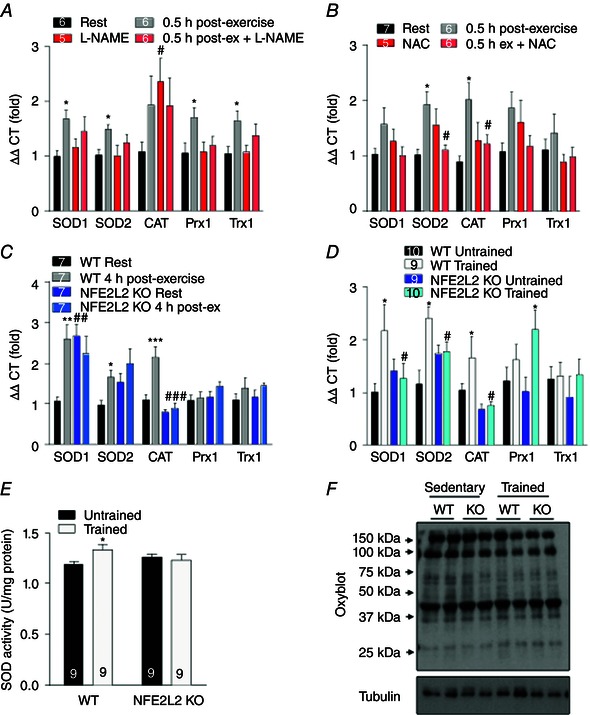
Key points: Reactive oxygen species (ROS) and nitric oxide (NO) regulate exercise-induced nuclear factor erythroid 2-related factor 2 (NFE2L2) expression in skeletal muscle. NFE2L2 is required for acute exercise-induced increases in skeletal muscle mitochondrial biogenesis genes, such as nuclear respiratory factor 1 (NRF-1) and mitochondrial transcription factor A, and anti-oxidant genes, such as superoxide dismutase (SOD)1, SOD2 and catalase. Following exercise training mice with impaired NFE2L2 expression have reduced exercise performance, energy expenditure, mitochondrial volume and anti-oxidant activity. In muscle cells, ROS and NO can regulate mitochondrial biogenesis via a NFE2L2/NRF-1-dependent pathway.
Abstract: Regular exercise induces adaptations to skeletal muscle, which can include mitochondrial biogenesis and enhanced anti-oxidant reserves. These adaptations and others are at least partly responsible for the improved health of physically active individuals. Reactive oxygen species (ROS) and nitric oxide (NO) are produced during exercise and may mediate the adaptive response to exercise in skeletal muscle. However, the mechanisms through which they act are unclear. In the present study, we aimed to determine the role of the redox-sensitive transcription factor nuclear factor erythroid-derived 2-like 2 (NFE2L2) in acute exercise- and training-induced mitochondrial biogenesis and the anti-oxidant response. We report that ROS and NO regulate acute exercise-induced expression of NFE2L2 in mouse skeletal muscle and muscle cells, and that deficiency in NFE2L2 prevents normal acute treadmill exercise-induced increases in mRNA of the mitochondrial biogenesis markers, nuclear respiratory factor 1 (NRF-1) and mitochondrial transcription factor A (mtTFA), and the anti-oxidants superoxide dismutase (SOD) 1 and 2, as well as catalase, in mouse gastrocnemius muscle. Furthermore, after 5 weeks of treadmill exercise training, mice deficient in NFE2L2 had reduced exercise capacity and whole body energy expenditure, as well as skeletal muscle mitochondrial mass and SOD activity, compared to wild-type littermates. In C2C12 myoblasts, acute treatment with exogenous H2 O2 (ROS)- and diethylenetriamine/NO adduct (NO donor) induced increases in mtTFA, which was prevented by small interfering RNA and short hairpin RNA knockdown of either NFE2L2 or NRF-1. Our results suggest that, during exercise, ROS and NO can act via NFE2L2 to functionally regulate skeletal muscle mitochondrial biogenesis and anti-oxidant defence gene expression.
© 2016 The Authors. The Journal of Physiology © 2016 The Physiological Society.
Figures
References
-
- Anderson EJ, Lustig ME, Boyle KE, Woodlief TL, Kane DA, Lin CT, Price JW, 3rd , Kang L, Rabinovitch PS, Szeto HH, Houmard JA, Cortright RN, Wasserman DH & Neufer PD (2009). Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J Clin Invest 119, 573–581. - PMC - PubMed
-
- Bassel‐Duby R & Olson EN (2006). Signaling pathways in skeletal muscle remodeling. Annu Rev Biochem 75, 19–37. - PubMed
-
- Chan P, Di Monte DA, Luo JJ, DeLanney LE, Irwin I & Langston JW (1994). Rapid ATP loss caused by methamphetamine in the mouse striatum: relationship between energy impairment and dopaminergic neurotoxicity. J Neurochem 62, 2484–2487. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous