

Review
doi: 10.1038/nrn.2016.27.
The neurogenetics of alternative splicing
Affiliations
- PMID: 27094079
- PMCID: PMC4861142
- DOI: 10.1038/nrn.2016.27
Item in Clipboard
Review
The neurogenetics of alternative splicing
Nat Rev Neurosci.
2016 May.
Abstract
Alternative precursor-mRNA splicing is a key mechanism for regulating gene expression in mammals and is controlled by specialized RNA-binding proteins. The misregulation of splicing is implicated in multiple neurological disorders. We describe recent mouse genetic studies of alternative splicing that reveal its critical role in both neuronal development and the function of mature neurons. We discuss the challenges in understanding the extensive genetic programmes controlled by proteins that regulate splicing, both during development and in the adult brain.
Figures
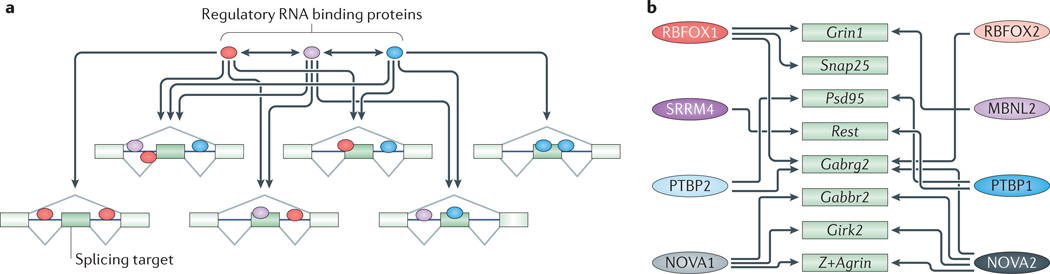
a | Splicing regulators control large target exon sets that often overlap with those regulated by other RNA-binding proteins (RBPs). Therefore, splicing of a given alternative exon can be affected by multiple RBPs. RBPs also affect their own splicing and homeostatic expression as well as that of other RBPs. The high degree of cross-regulation (indicated by arrows) between splicing regulators and their target sets creates complex splicing networks where the perturbation of a single RBP can lead to pleiotropic effects. Conversely, the splicing outcome of an exon can result from the combinatorial control of many RBPs. b | This figure shows some of the splicing target transcripts discussed in this Review (green boxes) that are cross-regulated (indicated by arrows) by multiple RBPs (coloured ovals on the left and right). Girk2, inwardly rectifying potassium channel Kir3.2; Gabrg2, GABAA receptor subunit gamma 2; Gabbr2, GABAB receptor 2; MBNL, muscleblind-like; NOVA, neuro-oncological ventral antigen; Psd95, postsynaptic density protein 95; PTBP, polypyrimidine tract binding protein; Rest, repressor element 1-silencing transcription factor; RBFOX, RNA-binding protein fox 1 homologue; Snap25, synaptosomal-associated protein 25; SRRM4, serine/arginine repetitive matrix protein 4.

Alternative splicing controls multiple aspects of early neuronal development. Defects in neurogenesis are seen in mouse mutants of a variety of regulatory RNA-binding proteins, including Ptbp1−/−, Ptbp2−/− and Srrm4−/−. Loss of polypyrimidine tract binding protein 1 (PTBP1) can cause precocious neurogenesis, deplete the neural stem cell pool and lead to fewer ependymal cells arising from radial glia later in development. PTBP2 loss may alter neural stem cell positioning and proliferation. Depletion of serine/arginine repetitive matrix protein 4 (SRRM4) inhibits neurogenesis of upper-layer neurons and causes the accumulation of progenitors or lower-layer neurons, resulting in abnormal cortical lamination (see inset). Defects in cortical lamination are also seen in mice lacking neuro-oncological ventral antigen 2 (NOVA2), where mis-splicing of the Reelin component disabled 1 (Dab1) leads to failure of many layer II/III and IV neurons to migrate properly (see inset). C P, cortical plate; IZ, intermediate zone; SVZ, subventricular zone; WM, white matter; VZ, ventricular zone. Figure (left panel) adapted with permission from REF. , Elsevier, and from REF. : Poduri, A., Evrony, G. D., Cai, X. & Walsh, C. A. Somatic mutation, genomic variation, and neurological disease. Science
341, 1237758–1237758 (2013). Reprinted with permission from AAAS.
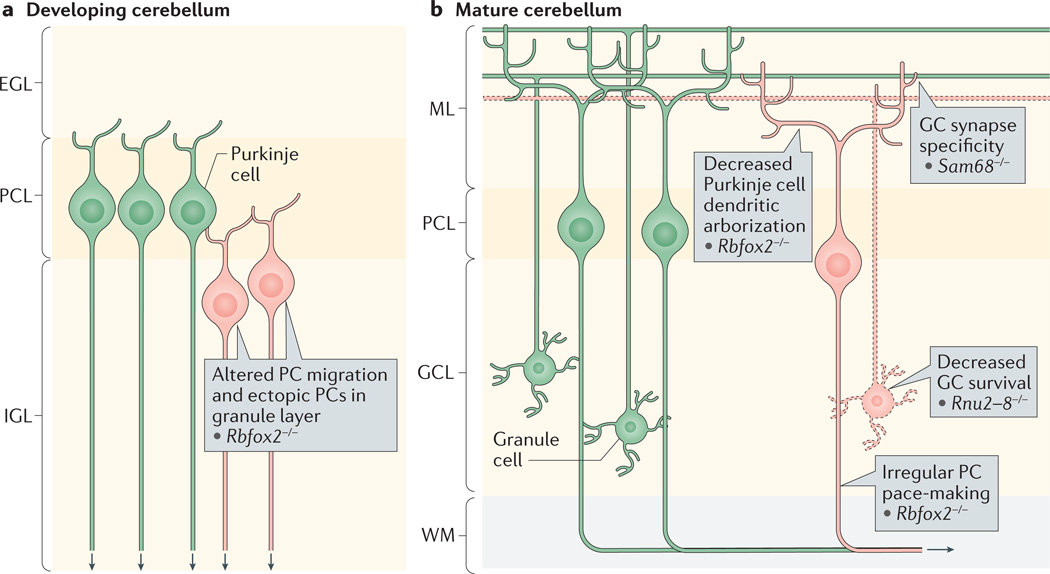
a | RNA-binding protein fox1 homologue 2 (RBFOX2) is required for both Purkinje cell (PC) migration and mature function. The cerebellum in Rbfox2−/− mouse mutants exhibits a disorganized PC layer (PCL) with ectopic PCs found in the internal granule layer (IGL), as well as reduced PC dendritic arborization later in development. b | In mature PCs, RBFOX2 controls the splicing and expression of the sodium channel gene Scn8a, which is needed for proper PC pace-making. Splicing regulation is also required for granule neuron survival and proper synaptic specificity. Loss of the U2 small nuclear RNA, a core spliceosomal component that is partially encoded by Rnu2–8, leads to increased intron retention and progressive granule neuron death. In granule neurons, SRC-associated in mitosis 68 kDa protein (SAM68) affects trans-synaptic interactions through alternative splicing of neurexin. EGL, external granule layer; GC, granule cell; GCL; granule cell layer; ML, molecular layer; WM, white matter.
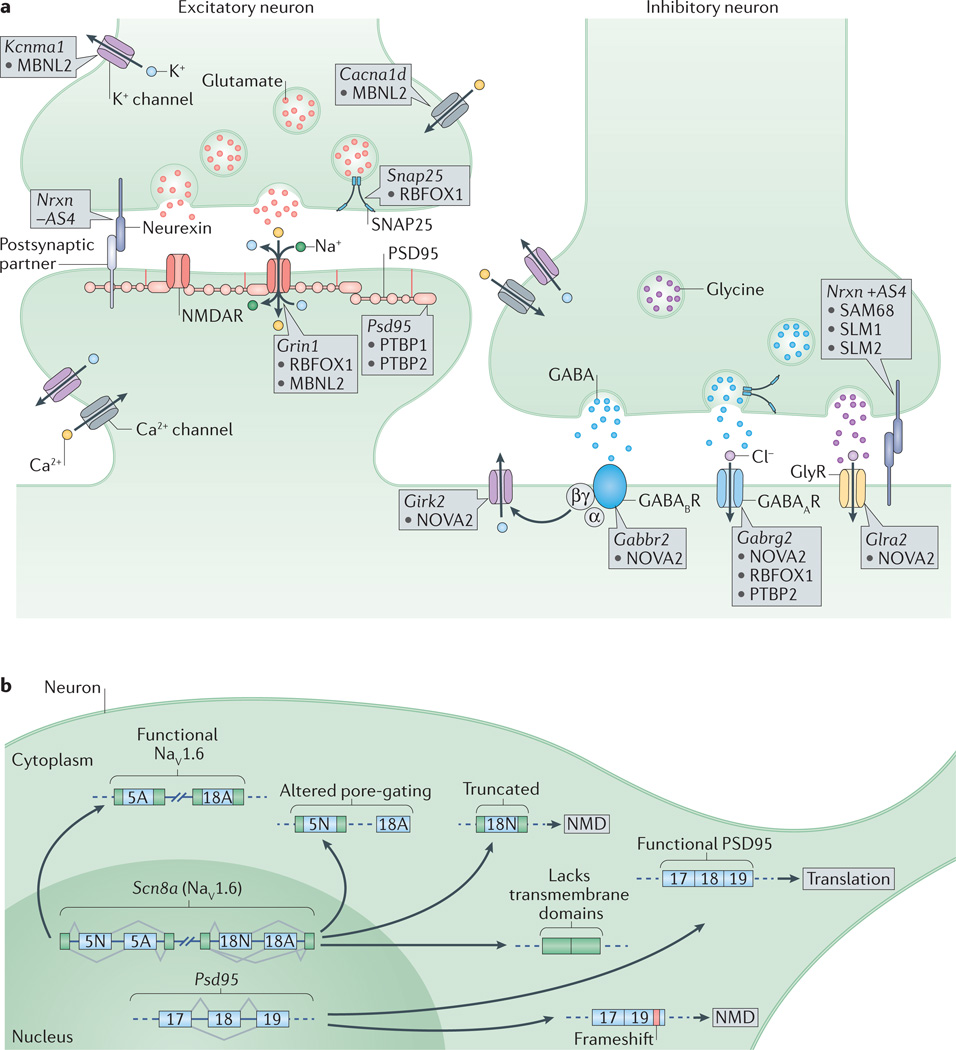
a |At the presynaptic terminal, alternative splicing of synaptosomal-associated protein 25 (Snap25) by RNA-binding protein fox 1 homologue 1 (RBFOX1) and of the calcium-activated potassium channel subunit alpha 1 (Kcnma1) by muscleblind-like 2 (MBNL2) are important to control neurotransmitter release. Differential splicing of the presynaptic neurexins (Nrxns) at AS4 by KHDRBS proteins (SAM68, SLM1 and SLM2) controls targeting to postsynaptic partners. At excitatory synapses, alternative splicing of the transcript encoding the NMDA receptor subunit GluN1, Grin1, is regulated by RBFOX1 and MBNL2, whereas the polypyrimidine tract binding proteins (PTBPs) control productive splicing of the scaffold protein, postsynaptic density protein 95 (Psd95). Splicing of the transcripts encoding L-type voltage-gated calcium channels, such as the pore-forming subunit Cav1.3 (encoded by Cacna1d), by MBNL2 may allow the voltage sensitivity, conductance, or other properties to be tuned as synapses differentiate. At inhibitory synapses, neuro-oncological ventral antigen 2 (NOVA2) mediates alternative splicing of the transcripts encoding many postsynaptic components such as the metabotropic GABAB receptor (Gabbr2), the inwardly rectifying potassium channel Kir3.2 (Girk2) and the glycine receptor alpha 2 (Glra2). Splicing of the GABAA receptor subunit transcript (Gabrg2) is controlled by multiple splicing regulators including NOVA2, RBFOX1 and PTBP2. b | Alternative splicing controls the expression and function of many synaptic components. Expression of PSD95 is repressed by PTBP-controlled exclusion of exon 18 until late in neuronal maturation when it is required for synaptogenesis. The gene encoding the voltage-gated sodium channel Nav1.6, Scn8a, has multiple alternative exons (such as 5N, 5A, 18N and 18A as shown in the figure) that can change its gating properties, determine its localization or alter its overall function. GABAAR, GABAA receptor; GABABR, GABAB receptor; GlyR, glycine receptor; NMD, nonsense-mediated decay; NMDAR, NMDA receptor; SAM68, SRC-associated in mitosis 68 kDa protein. Figure adapted from REF. , Nature Publishing Group.

RNA-binding proteins (RBPs) regulate transcript splicing, stability, translation and localization, and many RBPs studied as splicing regulators have extended functions in the cytoplasm, affecting every subsequent step of the mRNA life cycle. a | Nuclear roles of RBPs. Step 1: RBPs control alternative splicing of precursor mRNA (pre-mRNA) to generate multiple isoforms that differ in functional activity, interactions with cofactors or post-translational modifications. Step 2: regulated intron retention targets transcript isoforms for degradation by nuclear surveillance mechanisms,, or by the introduction of a premature termination codon leading to nonsense-mediated decay (NMD, shown in red; see below). Step 3: by shifting the reading frame or by including a ‘poison exon’ containing a premature translation termination codon, alternative splicing produces transcript isoforms that are degraded by NMD. Alternative splicing coupled with NMD can control the overall abundance of gene transcripts. b | Cytoplasmic roles of RBPs. Step 4: RBPs compete with AU-binding proteins for binding at AU-rich elements in the 3′ untranslated region (UTR) to stabilize their target transcripts,. Other RBPs regulate transcript stability in the cytoplasm by either competing with microRNAs for their binding sites or facilitating microRNA binding,,. RBP binding in both the 5′ UTRs and 3′ UTRs also affects translational efficiency. Step 5: RBPs regulate the transport and differential localization of mRNA, which are crucial for spatial and temporal control of translation in response to activity-dependent signalling,,. The establishment of neuronal polarity and consolidation of synaptic strength through local translation of mRNAs in response to synaptic activity are some well-known examples. AAA, poly-A tail; Gppp, 5′ cap; Pol II, RNA polymerase II.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
