

Function of the Borrelia burgdorferi FtsH Homolog Is Essential for Viability both In Vitro and In Vivo and Independent of HflK/C
- PMID: 27094329
- PMCID: PMC4850261
- DOI: 10.1128/mBio.00404-16
Function of the Borrelia burgdorferi FtsH Homolog Is Essential for Viability both In Vitro and In Vivo and Independent of HflK/C
Erratum in
-
Erratum for Chu et al., Function of the Borrelia burgdorferi FtsH Homolog Is Essential for Viability both In Vitro and In Vivo and Independent of HflK/C.mBio. 2016 Dec 20;7(6):e02135-16. doi: 10.1128/mBio.02135-16. mBio. 2016. PMID: 27999167 Free PMC article. No abstract available.
Abstract
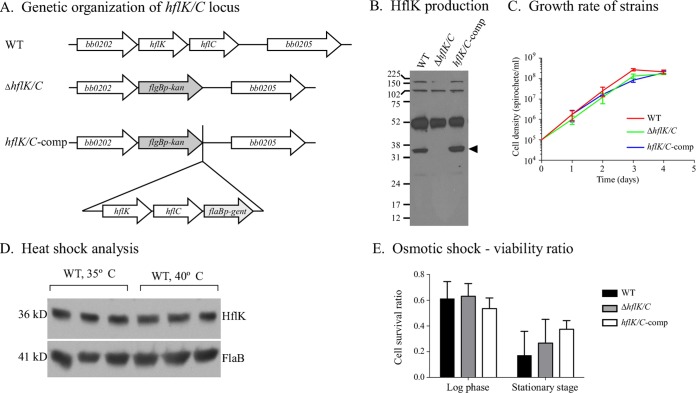
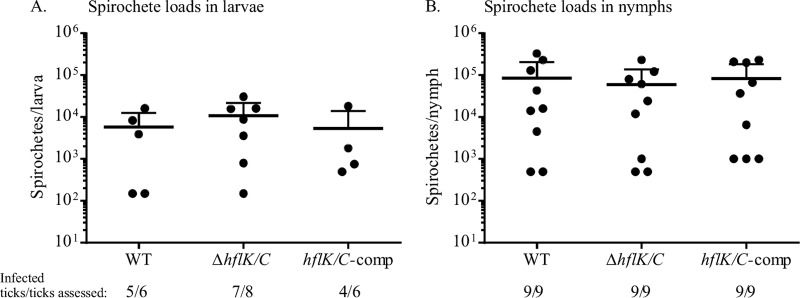
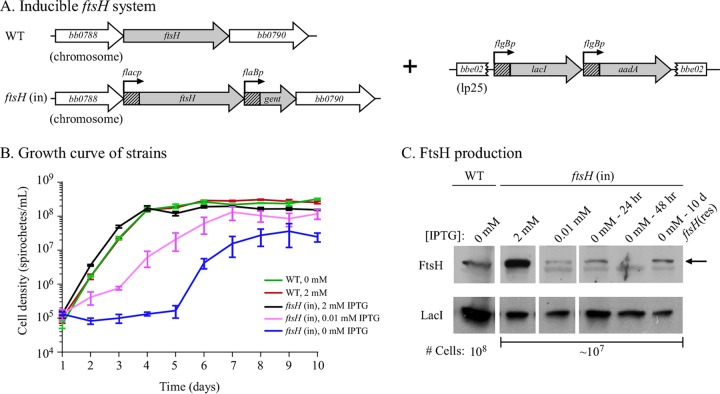
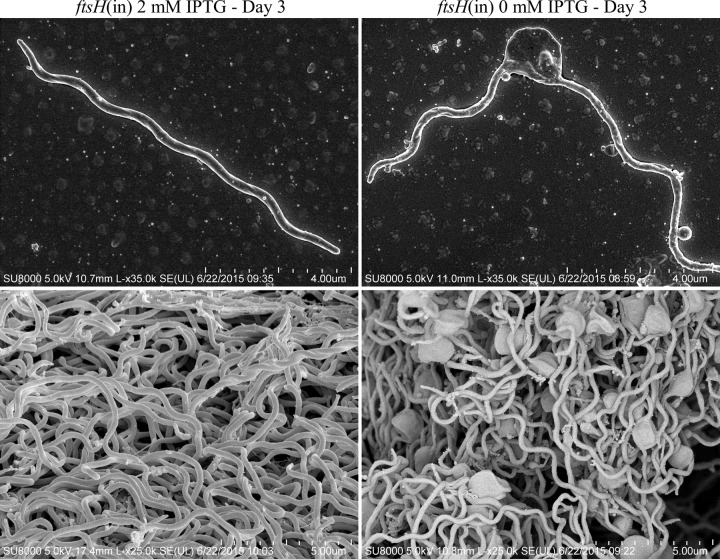
In many bacteria, the FtsH protease and its modulators, HflK and HflC, form a large protein complex that contributes to both membrane protein quality control and regulation of the cellular response to environmental stress. Both activities are crucial to the Lyme disease pathogen Borrelia burgdorferi, which depends on membrane functions, such as motility, protein transport, and cell signaling, to respond to rapid changes in its environment. Using an inducible system, we demonstrate that FtsH production is essential for both mouse and tick infectivity and for in vitro growth of B. burgdorferi FtsH depletion in B. burgdorferi cells resulted in membrane deformation and cell death. Overproduction of the protease did not have any detectable adverse effects on B. burgdorferi growth in vitro, suggesting that excess FtsH does not proteolytically overwhelm its substrates. In contrast, we did not observe any phenotype for cells lacking the protease modulators HflK and HflC (ΔHflK/C), although we examined morphology, growth rate, growth under stress conditions, and the complete mouse-tick infectious cycle. Our results demonstrate that FtsH provides an essential function in the life cycle of the obligate pathogen B. burgdorferi but that HflK and HflC do not detectably affect FtsH function.
Importance: Lyme disease is caused by Borrelia burgdorferi, which is maintained in nature in an infectious cycle alternating between small mammals and Ixodes ticks. B. burgdorferi produces specific membrane proteins to successfully infect and persist in these diverse organisms. We hypothesized that B. burgdorferi has a specific mechanism to ensure that membrane proteins are properly folded and biologically active when needed and removed if improperly folded or dysfunctional. Our experiments demonstrate that FtsH, a protease that fulfills this role in other microorganisms, is essential to B. burgdorferi viability. Cells depleted of FtsH do not survive in laboratory culture medium and cannot colonize mice or ticks, revealing an absolute requirement for this protease. However, the loss of two potential modulators of FtsH activity, HflK and HflC, does not detectably affect B. burgdorferi physiology. Our results provide the groundwork for the identification of FtsH substrates that are critical for the bacterium's viability.
Copyright © 2016 Chu et al.
Figures
Similar articles
-
Structure of the Borrelia burgdorferi ATP-dependent metalloprotease FtsH in its functionally relevant hexameric form.Biochim Biophys Acta Proteins Proteom. 2024 Jan 1;1872(1):140969. doi: 10.1016/j.bbapap.2023.140969. Epub 2023 Oct 16. Biochim Biophys Acta Proteins Proteom. 2024. PMID: 37852516
-
Stage-specific global alterations in the transcriptomes of Lyme disease spirochetes during tick feeding and following mammalian host adaptation.Mol Microbiol. 2015 Feb;95(3):509-38. doi: 10.1111/mmi.12882. Epub 2014 Dec 30. Mol Microbiol. 2015. PMID: 25425211 Free PMC article.
-
Long-term survival of Borrelia burgdorferi lacking the hibernation promotion factor homolog in the unfed tick vector.Infect Immun. 2015 Dec;83(12):4800-10. doi: 10.1128/IAI.00925-15. Epub 2015 Oct 5. Infect Immun. 2015. PMID: 26438790 Free PMC article.
-
Physiologic and Genetic Factors Influencing the Zoonotic Cycle of Borrelia burgdorferi.Curr Top Microbiol Immunol. 2018;415:63-82. doi: 10.1007/82_2017_43. Curr Top Microbiol Immunol. 2018. PMID: 28864829 Review.
-
The Consistent Tick-Vertebrate Infectious Cycle of the Lyme Disease Spirochete Enables Borrelia burgdorferi To Control Protein Expression by Monitoring Its Physiological Status.J Bacteriol. 2022 May 17;204(5):e0060621. doi: 10.1128/jb.00606-21. Epub 2022 Apr 5. J Bacteriol. 2022. PMID: 35380872 Free PMC article. Review.
Cited by
-
Lyme disease: the promise of Big Data, companion diagnostics and precision medicine.Infect Drug Resist. 2016 Sep 13;9:215-9. doi: 10.2147/IDR.S114770. eCollection 2016. Infect Drug Resist. 2016. PMID: 27672336 Free PMC article.
-
Two FtsH Proteases Contribute to Fitness and Adaptation of Pseudomonas aeruginosa Clone C Strains.Front Microbiol. 2019 Jul 9;10:1372. doi: 10.3389/fmicb.2019.01372. eCollection 2019. Front Microbiol. 2019. PMID: 31338071 Free PMC article.
-
The Lon-1 Protease Is Required by Borrelia burgdorferi To Infect the Mammalian Host.Infect Immun. 2020 May 20;88(6):e00951-19. doi: 10.1128/IAI.00951-19. Print 2020 May 20. Infect Immun. 2020. PMID: 32205400 Free PMC article.
-
Dissection of amino acid acquisition pathways in Borrelia burgdorferi uncovers unique physiological responses.bioRxiv [Preprint]. 2025 Mar 15:2025.03.14.643351. doi: 10.1101/2025.03.14.643351. bioRxiv. 2025. PMID: 40161780 Free PMC article. Preprint.
-
Coupled induction of prophage and virulence factors during tick transmission of the Lyme disease spirochete.Nat Commun. 2023 Jan 13;14(1):198. doi: 10.1038/s41467-023-35897-3. Nat Commun. 2023. PMID: 36639656 Free PMC article.
References
-
- Ogura T, Inoue K, Tatsuta T, Suzaki T, Karata K, Young K, Su LH, Fierke CA, Jackman JE, Raetz CR, Coleman J, Tomoyasu T, Matsuzawa H. 1999. Balanced biosynthesis of major membrane components through regulated degradation of the committed enzyme of lipid A biosynthesis by the AAA protease FtsH (HflB) in Escherichia coli. Mol Microbiol 31:833–844. doi: 10.1046/j.1365-2958.1999.01221.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
