

Interferon-gamma potentiates NMDA receptor signaling in spinal dorsal horn neurons via microglia-neuron interaction
- PMID: 27094552
- PMCID: PMC4956380
- DOI: 10.1177/1744806916644927
Interferon-gamma potentiates NMDA receptor signaling in spinal dorsal horn neurons via microglia-neuron interaction
Abstract
Background: Glia-neuron interactions play an important role in the development of neuropathic pain. Expression of the pro-inflammatory cytokne →cytokine Interferon-gamma (IFNγ) is upregulated in the dorsal horn after peripheral nerve injury, and intrathecal IFNγ administration induces mechanical allodynia in rats. A growing body of evidence suggests that IFNγ might be involved in the mechanisms of neuropathic pain, but its effects on the spinal dorsal horn are unclear. We performed blind whole-cell patch-clamp recording to investigate the effect of IFNγ on postsynaptic glutamate-induced currents in the substantia gelatinosa neurons of spinal cord slices from adult male rats.
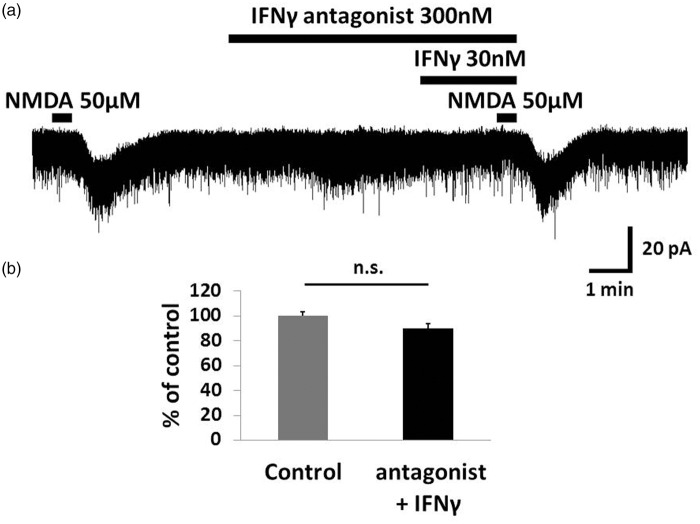
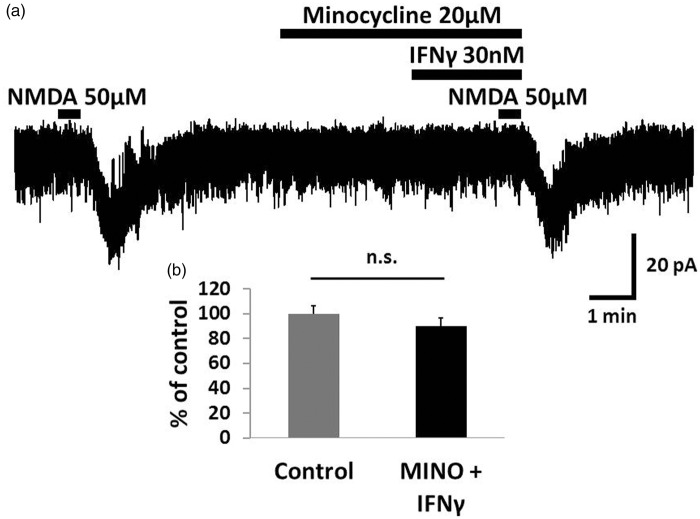
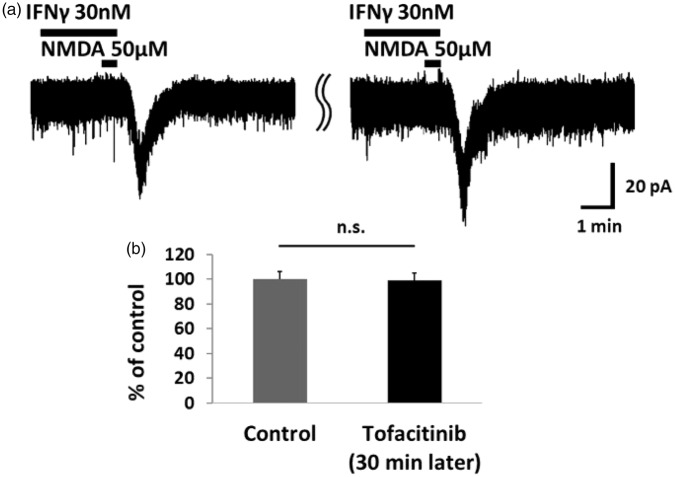
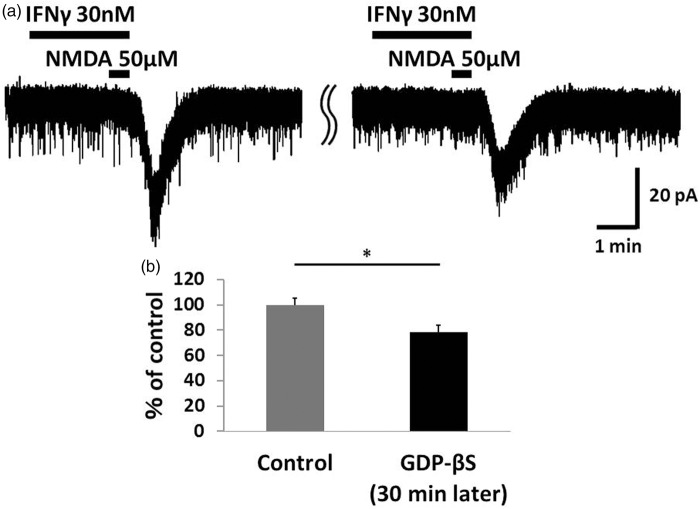
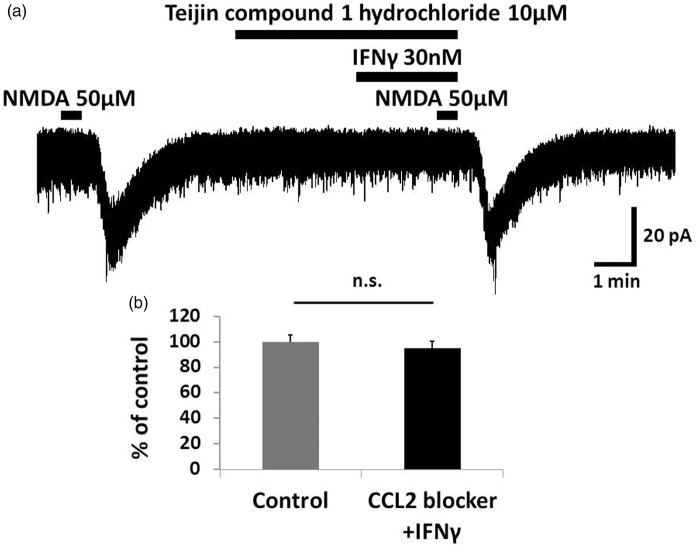
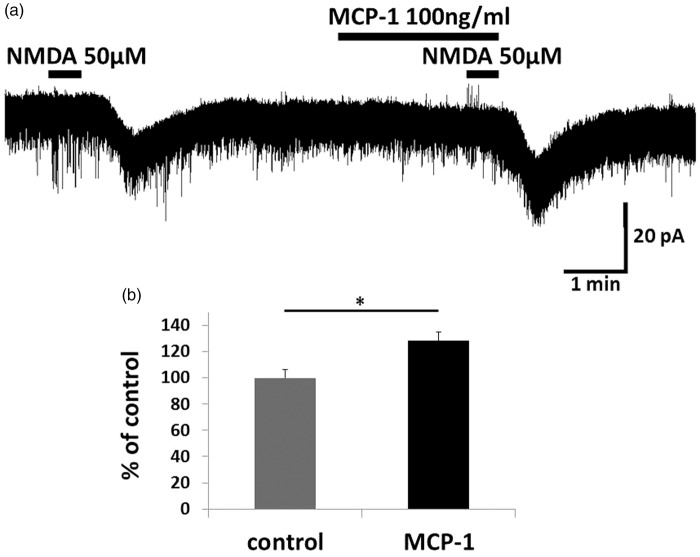
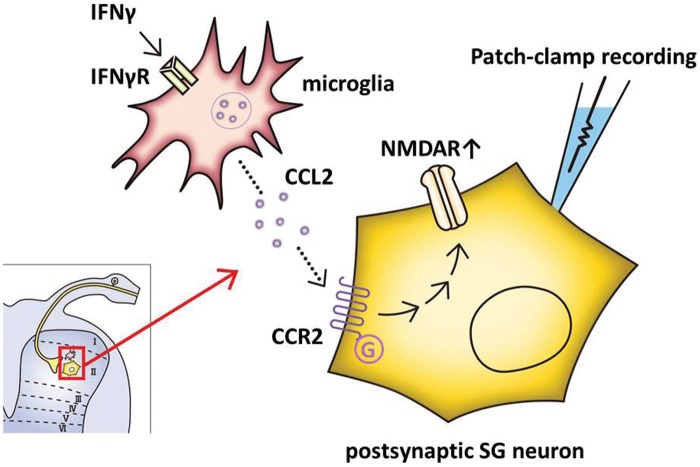
Results: IFNγ perfusion significantly enhanced the amplitude of NMDA-induced inward currents in substantia gelatinosa neurons, but did not affect AMPA-induced currents. The facilitation of NMDA-induced current by IFNγ was inhibited by bath application of an IFNγ receptor-selective antagonist. Adding the Janus activated kinase inhibitor tofacitinib to the pipette solution did not affect the IFNγ-induced facilitation of NMDA-induced currents. However, the facilitatory effect of IFNγ on NMDA-induced currents was inhibited by perfusion of the microglial inhibitor minocycline. These results suggest that IFNγ binds the microglial IFNγ receptor and enhances NMDA receptor activity in substantia gelatinosa neurons. Next, to identify the effector of signal transmission from microglia to dorsal horn neurons, we added an inhibitor of G proteins, GDP-β-S, to the pipette solution. In a GDP-β-S-containing pipette solution, IFNγ-induced potentiation of the NMDA current was significantly suppressed after 30 min. In addition, IFNγ-induced potentiation of NMDA currents was blocked by application of a selective antagonist of CCR2, and its ligand CCL2 increased NMDA-induced currents.
Conclusion: Our findings suggest that IFNγ enhance the amplitude of NMDA-induced inward currents in substantia gelatinosa neurons via microglial IFNγ receptors and CCL2/CCR2 signaling. This mechanism might be partially responsible for the development of persistent neuropathic pain.
Keywords: CCR2; NMDA receptor; Neuropathic pain; dorsal horn; interferon-gamma; microglia; patch-clamp recording; spinal cord.
© The Author(s) 2016.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
