

Virtual memory T cells develop and mediate bystander protective immunity in an IL-15-dependent manner
- PMID: 27097762
- PMCID: PMC4844673
- DOI: 10.1038/ncomms11291
Virtual memory T cells develop and mediate bystander protective immunity in an IL-15-dependent manner
Abstract
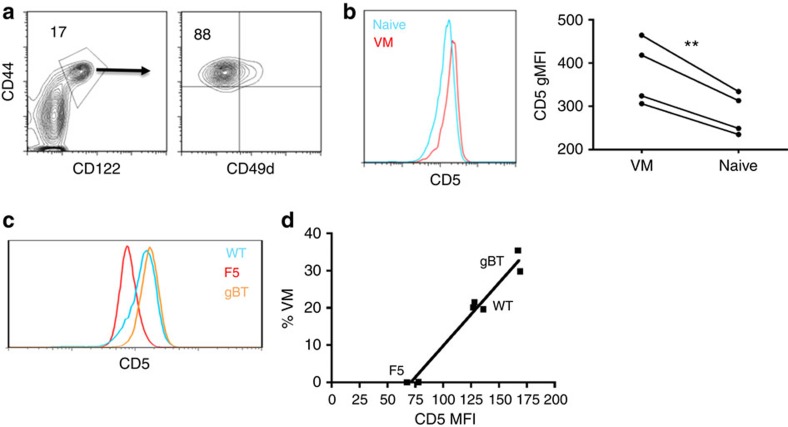
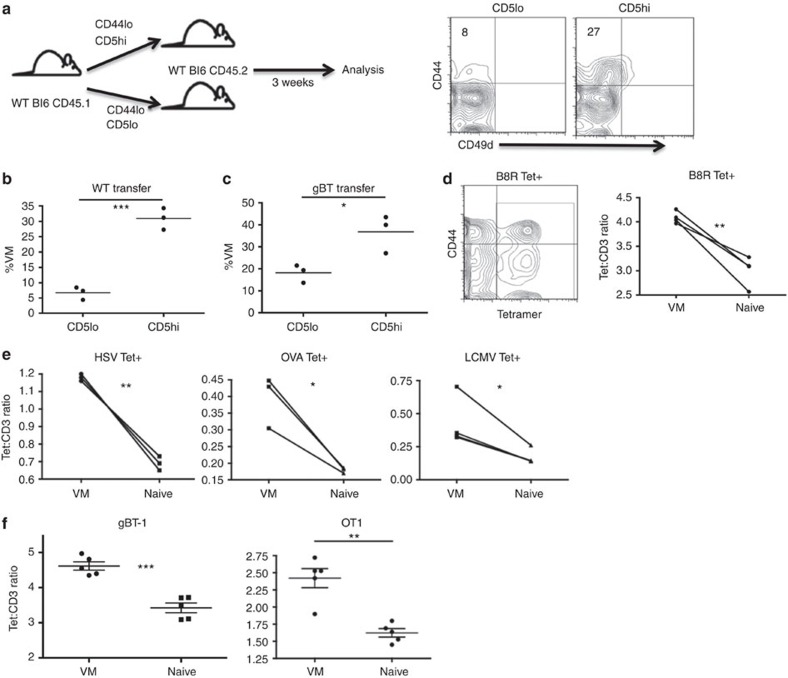
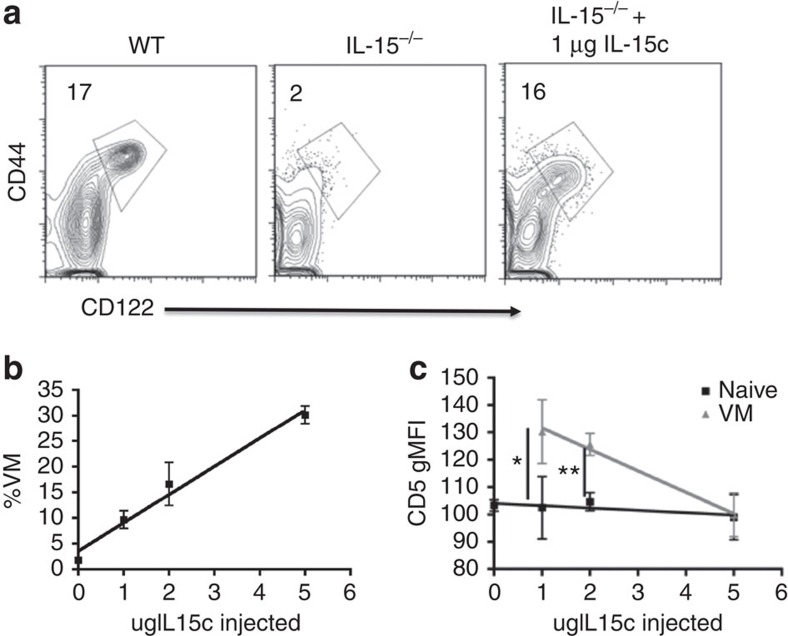
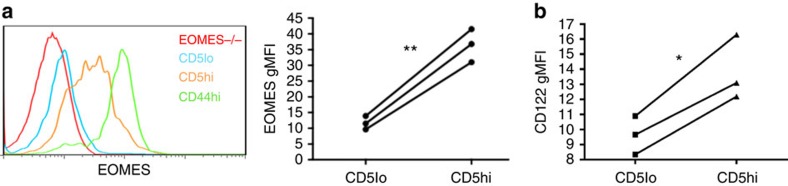
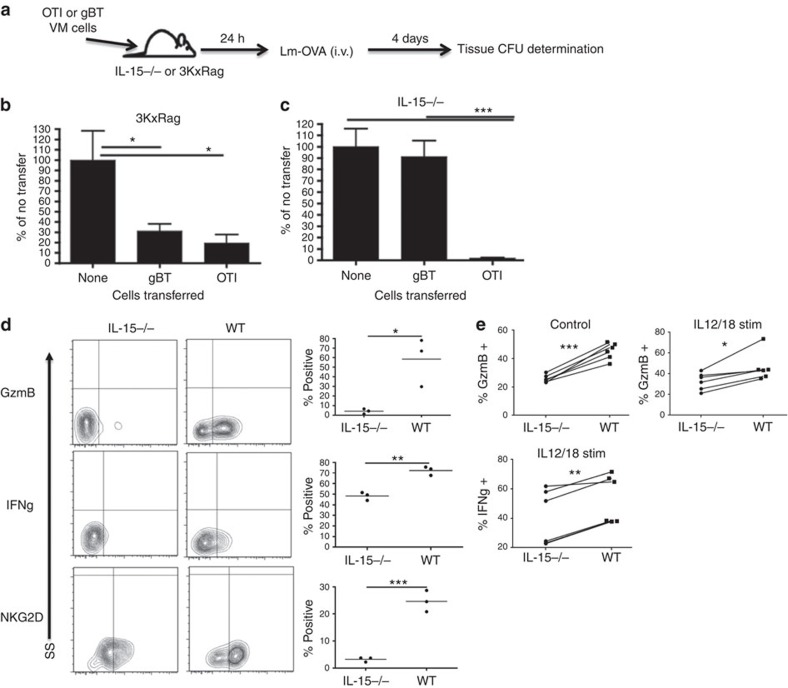
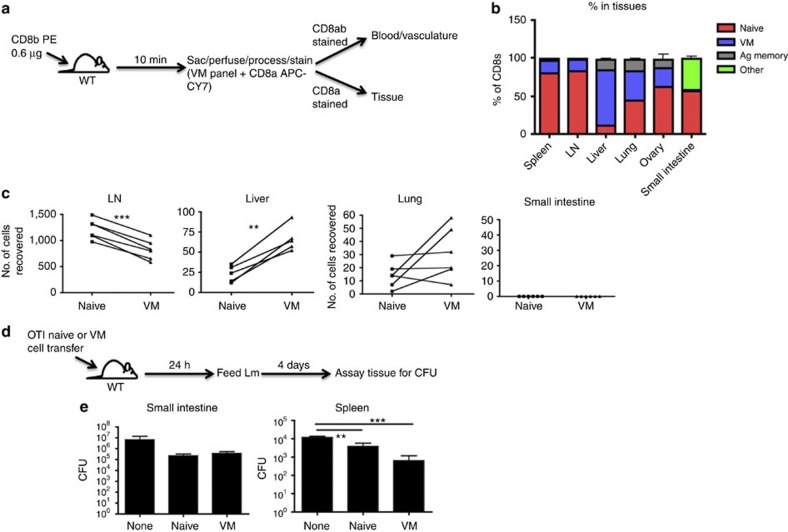
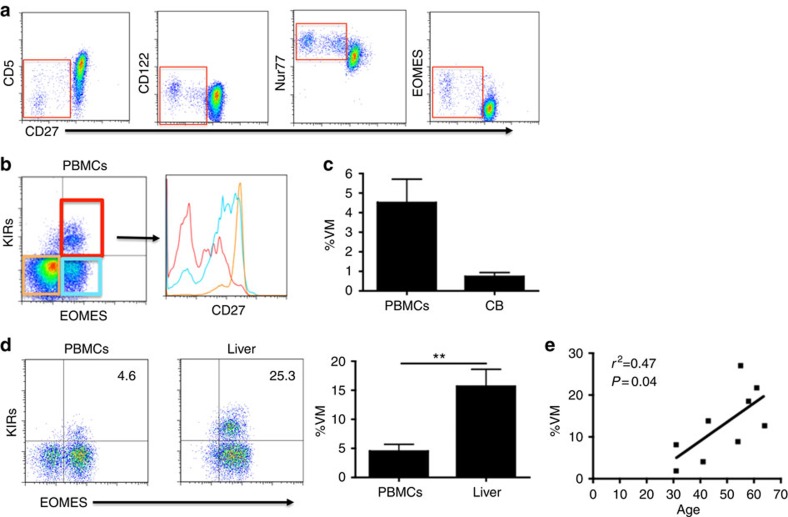
Virtual memory cells (VM) are an antigen-specific, memory phenotype CD8 T-cell subset found in lymphoreplete, unchallenged mice. Previous studies indicated that VM cells were the result of homeostatic proliferation (HP) resembling the proliferation observed in a lymphopenic environment. Here we demonstrate that HP is ongoing in lymphoreplete mice, the degree of which is dictated by the number of naive CD8 T cells with a sufficiently high affinity for self-antigen interacting with peripheral IL-15. VM cell transcriptional profiles suggest a capacity to mediate protective immunity via antigen non-specific bystander killing, a function we show is dependent on IL-15. Finally, we show a VM-like population of human cells that accumulate with age and traffic to the liver, displaying phenotypic and functional attributes consistent with the bystander protective functions of VM cells identified in the mouse. These data identify developmental and functional attributes of VM cells, including their likely role in protective immunity.
Figures
References
-
- Huang T., Wei B., Velazquez P., Borneman J. & Braun J. Commensal microbiota alter the abundance and TCR responsiveness of splenic naive CD4+ T lymphocytes. Clin. Immunol. 117, 221–230 (2005) . - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
