

Coupling switch of P2Y-IP3 receptors mediates differential Ca(2+) signaling in human embryonic stem cells and derived cardiovascular progenitor cells
- PMID: 27098757
- PMCID: PMC5023627
- DOI: 10.1007/s11302-016-9512-9
Coupling switch of P2Y-IP3 receptors mediates differential Ca(2+) signaling in human embryonic stem cells and derived cardiovascular progenitor cells
Abstract
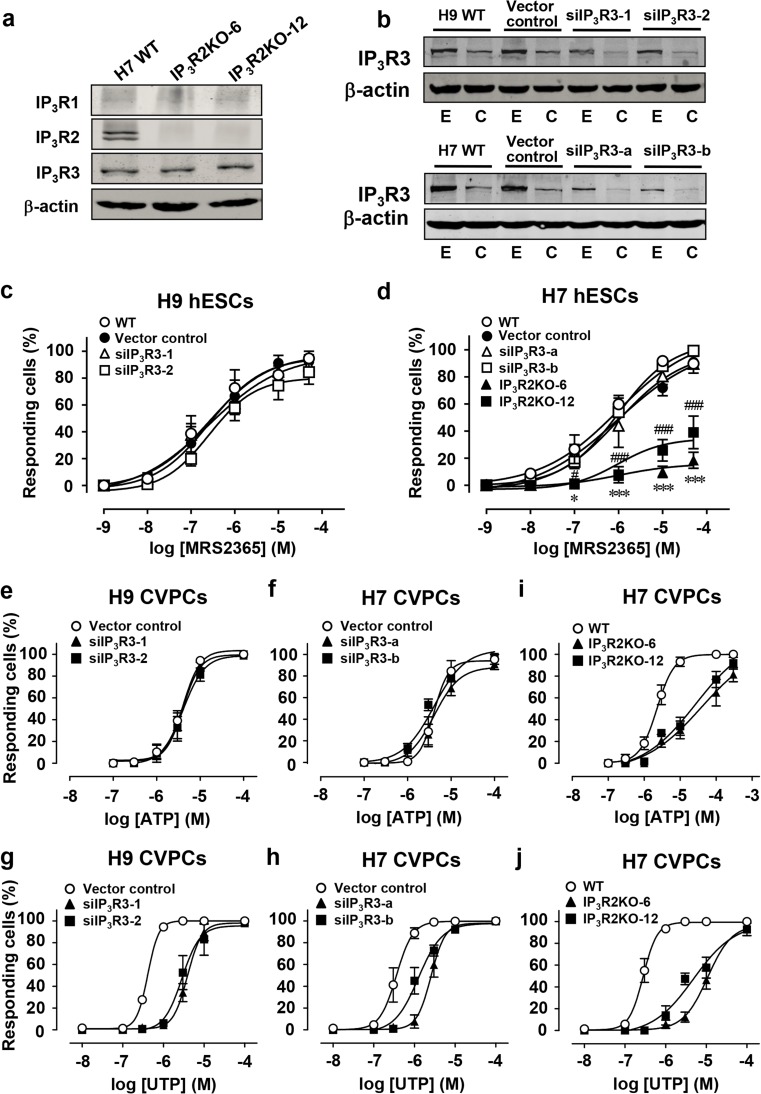
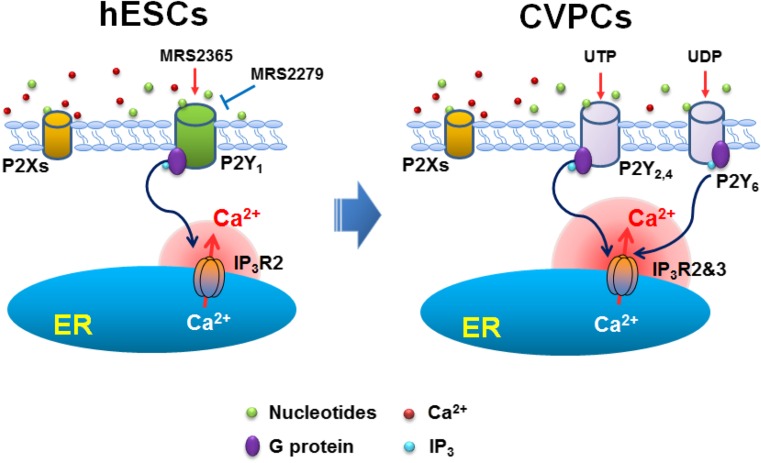
Purinergic signaling mediated by P2 receptors (P2Rs) plays important roles in embryonic and stem cell development. However, how it mediates Ca(2+) signals in human embryonic stem cells (hESCs) and derived cardiovascular progenitor cells (CVPCs) remains unclear. Here, we aimed to determine the role of P2Rs in mediating Ca(2+) mobilizations of these cells. hESCs were induced to differentiate into CVPCs by our recently established methods. Gene expression of P2Rs and inositol 1,4,5-trisphosphate receptors (IP3Rs) was analyzed by quantitative/RT-PCR. IP3R3 knockdown (KD) or IP3R2 knockout (KO) hESCs were established by shRNA- or TALEN-mediated gene manipulations, respectively. Confocal imaging revealed that Ca(2+) responses in CVPCs to ATP and UTP were more sensitive and stronger than those in hESCs. Consistently, the gene expression levels of most P2YRs except P2Y1 were increased in CVPCs. Suramin or PPADS blocked ATP-induced Ca(2+) transients in hESCs but only partially inhibited those in CVPCs. Moreover, the P2Y1 receptor-specific antagonist MRS2279 abolished most ATP-induced Ca(2+) signals in hESCs but not in CVPCs. P2Y1 receptor-specific agonist MRS2365 induced Ca(2+) transients only in hESCs but not in CVPCs. Furthermore, IP3R2KO but not IP3R3KD decreased the proportion of hESCs responding to MRS2365. In contrast, both IP3R2 and IP3R3 contributed to UTP-induced Ca(2+) responses while ATP-induced Ca(2+) responses were more dependent on IP3R2 in the CVPCs. In conclusion, a predominant role of P2Y1 receptors in hESCs and a transition of P2Y-IP3R coupling in derived CVPCs are responsible for the differential Ca(2+) mobilization between these cells.
Keywords: Ca2+ signaling; IP3 receptors; Lineage progenitors; P2Y receptors; Pluripotent stem cells.
Conflict of interest statement
The authors declare that they have no conflicts of interest.
Figures






Similar articles
-
Minimal contribution of IP3R2 in cardiac differentiation and derived ventricular-like myocytes from human embryonic stem cells.Acta Pharmacol Sin. 2020 Dec;41(12):1576-1586. doi: 10.1038/s41401-020-00528-w. Epub 2020 Oct 9. Acta Pharmacol Sin. 2020. PMID: 33037404 Free PMC article.
-
ATP induces contraction of cultured brain capillary pericytes via activation of P2Y-type purinergic receptors.Am J Physiol Heart Circ Physiol. 2021 Feb 1;320(2):H699-H712. doi: 10.1152/ajpheart.00560.2020. Epub 2020 Dec 11. Am J Physiol Heart Circ Physiol. 2021. PMID: 33306443
-
Expression and reconstitution of the bioluminescent Ca(2+) reporter aequorin in human embryonic stem cells, and exploration of the presence of functional IP3 and ryanodine receptors during the early stages of their differentiation into cardiomyocytes.Sci China Life Sci. 2016 Aug;59(8):811-24. doi: 10.1007/s11427-016-5094-6. Epub 2016 Jul 19. Sci China Life Sci. 2016. PMID: 27430888
-
Inositol 1,4,5-trisphosphate receptor-isoform diversity in cell death and survival.Biochim Biophys Acta. 2014 Oct;1843(10):2164-83. doi: 10.1016/j.bbamcr.2014.03.007. Epub 2014 Mar 15. Biochim Biophys Acta. 2014. PMID: 24642269 Review.
-
Molecular pharmacology of P2Y-receptors.Naunyn Schmiedebergs Arch Pharmacol. 2000 Nov;362(4-5):310-23. doi: 10.1007/s002100000310. Naunyn Schmiedebergs Arch Pharmacol. 2000. PMID: 11111826 Review.
Cited by
-
Cardiac Na+-Ca2+ exchanger 1 (ncx1h) is critical for the ventricular cardiomyocyte formation via regulating the expression levels of gata4 and hand2 in zebrafish.Sci China Life Sci. 2021 Feb;64(2):255-268. doi: 10.1007/s11427-019-1706-1. Epub 2020 Jul 7. Sci China Life Sci. 2021. PMID: 32648190
-
Extracellular vesicles from human embryonic stem cell-derived cardiovascular progenitor cells promote cardiac infarct healing through reducing cardiomyocyte death and promoting angiogenesis.Cell Death Dis. 2020 May 11;11(5):354. doi: 10.1038/s41419-020-2508-y. Cell Death Dis. 2020. PMID: 32393784 Free PMC article.
-
Functional expression of the Ca2+ signaling machinery in human embryonic stem cells.Acta Pharmacol Sin. 2017 Dec;38(12):1663-1672. doi: 10.1038/aps.2017.29. Epub 2017 Jul 10. Acta Pharmacol Sin. 2017. PMID: 28713161 Free PMC article.
-
TATA box-binding protein-related factor 3 drives the mesendoderm specification of human embryonic stem cells by globally interacting with the TATA box of key mesendodermal genes.Stem Cell Res Ther. 2020 May 24;11(1):196. doi: 10.1186/s13287-020-01711-w. Stem Cell Res Ther. 2020. PMID: 32448362 Free PMC article.
-
Myosin light chain 2 marks differentiating ventricular cardiomyocytes derived from human embryonic stem cells.Pflugers Arch. 2021 Jul;473(7):991-1007. doi: 10.1007/s00424-021-02578-3. Epub 2021 May 24. Pflugers Arch. 2021. PMID: 34031754
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
