

Neutralization of Virus Infectivity by Antibodies: Old Problems in New Perspectives
- PMID: 27099867
- PMCID: PMC4835181
- DOI: 10.1155/2014/157895
Neutralization of Virus Infectivity by Antibodies: Old Problems in New Perspectives
Abstract
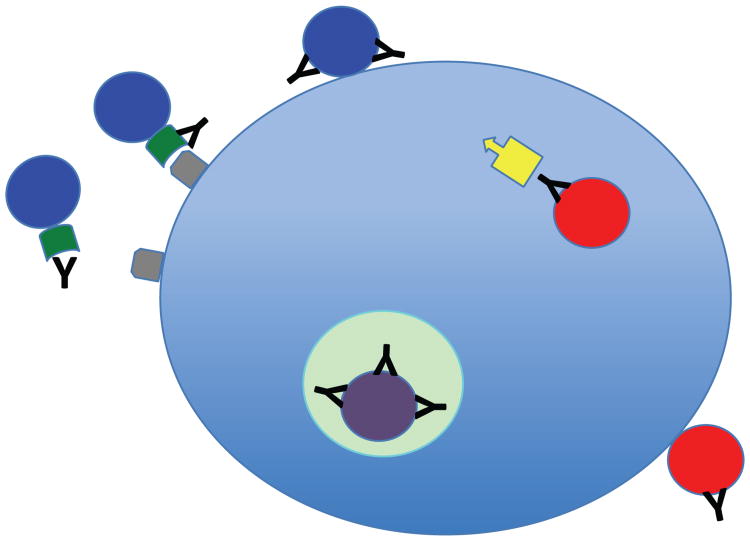
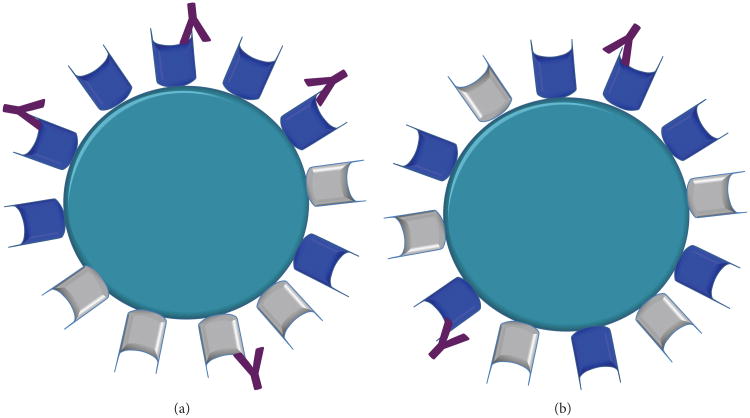
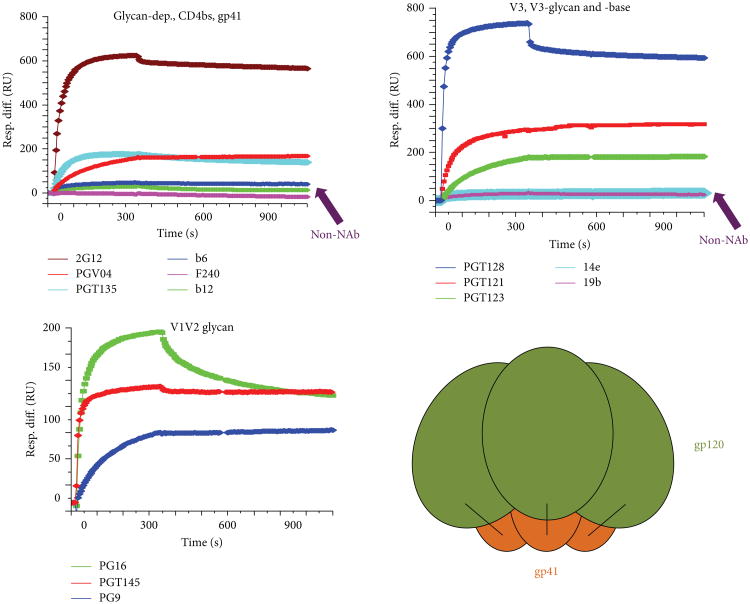
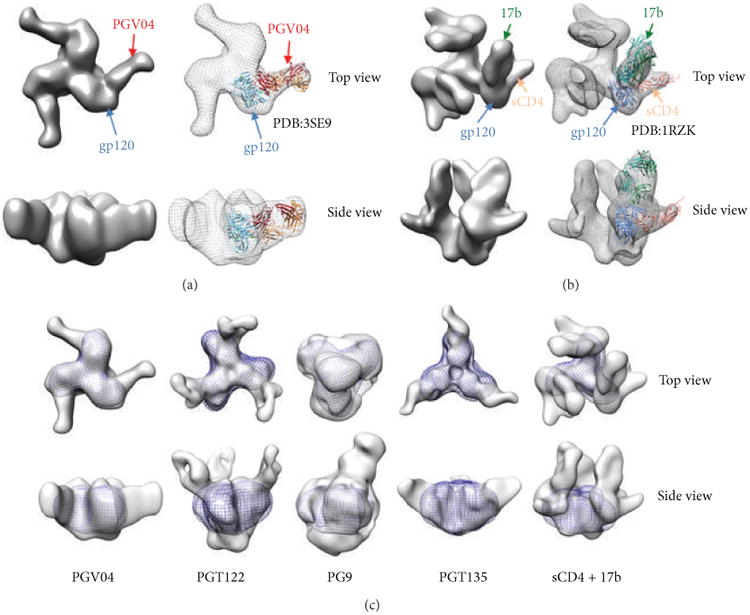
Neutralizing antibodies (NAbs) can be both sufficient and necessary for protection against viral infections, although they sometimes act in concert with cellular immunity. Successful vaccines against viruses induce NAbs but vaccine candidates against some major viral pathogens, including HIV-1, have failed to induce potent and effective such responses. Theories of how antibodies neutralize virus infectivity have been formulated and experimentally tested since the 1930s; and controversies about the mechanistic and quantitative bases for neutralization have continually arisen. Soluble versions of native oligomeric viral proteins that mimic the functional targets of neutralizing antibodies now allow the measurement of the relevant affinities of NAbs. Thereby the neutralizing occupancies on virions can be estimated and related to the potency of the NAbs. Furthermore, the kinetics and stoichiometry of NAb binding can be compared with neutralizing efficacy. Recently, the fundamental discovery that the intracellular factor TRIM21 determines the degree of neutralization of adenovirus has provided new mechanistic and quantitative insights. Since TRIM21 resides in the cytoplasm, it would not affect the neutralization of enveloped viruses, but its range of activity against naked viruses will be important to uncover. These developments bring together the old problems of virus neutralization-mechanism, stoichiometry, kinetics, and efficacy-from surprising new angles.
Conflict of interest statement
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources