

Musculocontractural Ehlers-Danlos syndrome and neurocristopathies: dermatan sulfate is required for Xenopus neural crest cells to migrate and adhere to fibronectin
- PMID: 27101845
- PMCID: PMC4920151
- DOI: 10.1242/dmm.024661
Musculocontractural Ehlers-Danlos syndrome and neurocristopathies: dermatan sulfate is required for Xenopus neural crest cells to migrate and adhere to fibronectin
Abstract
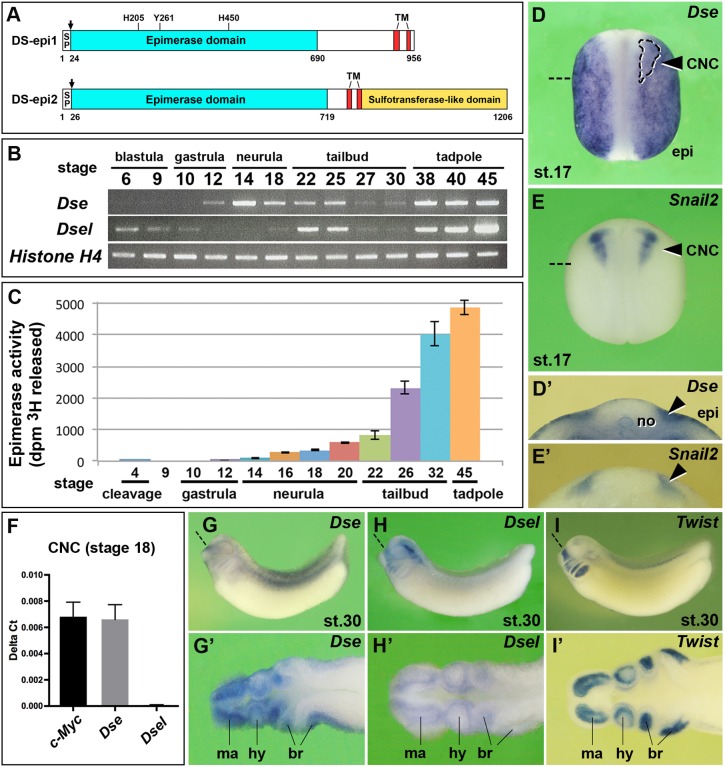
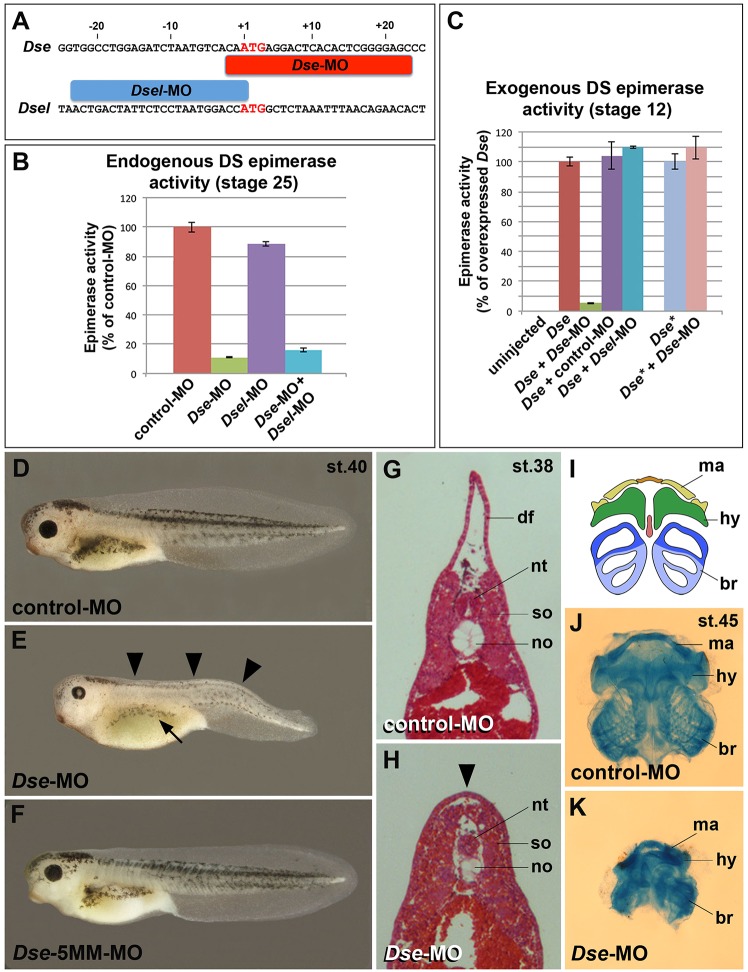
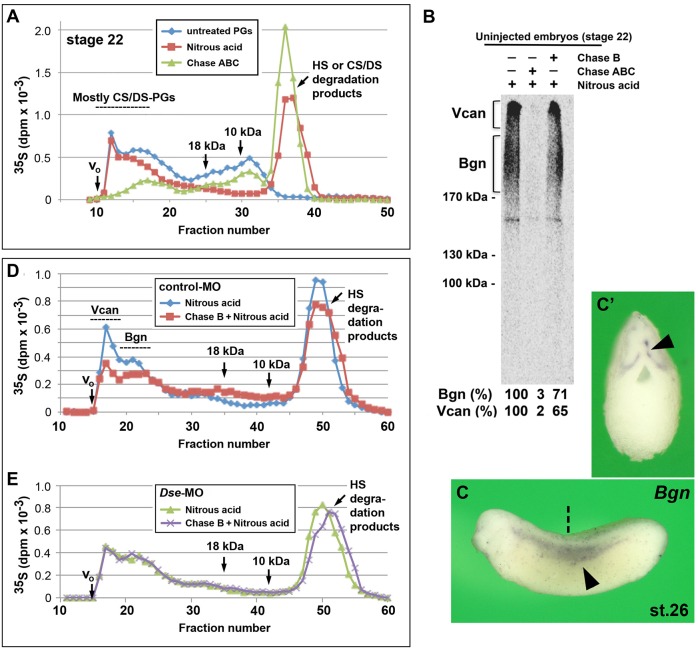
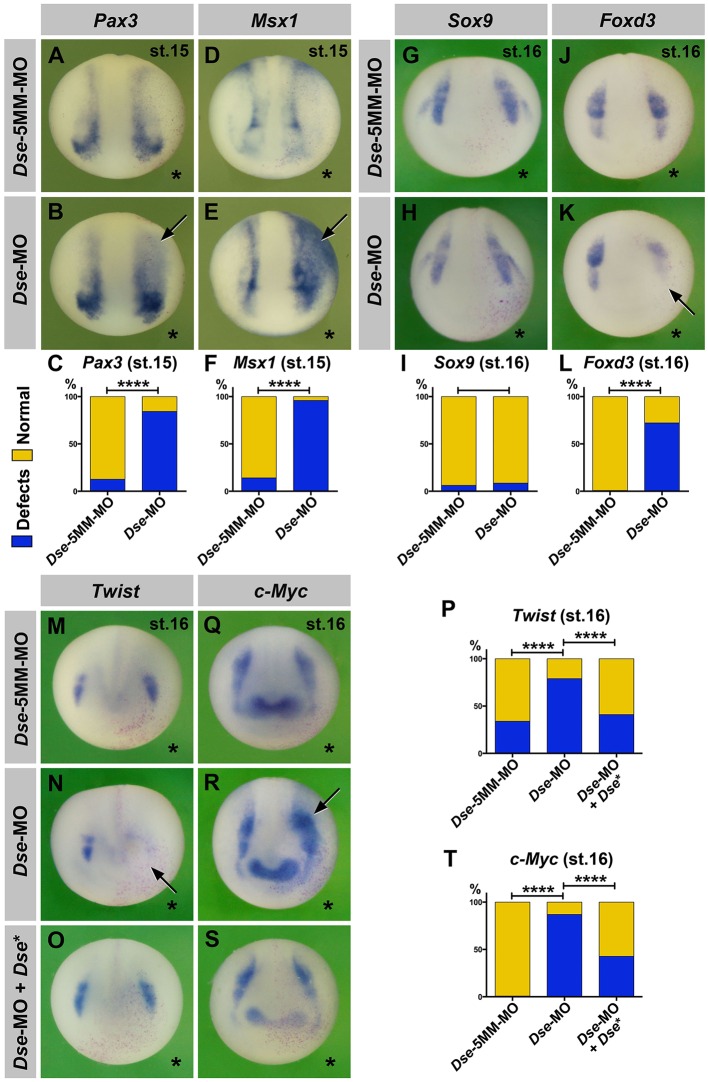
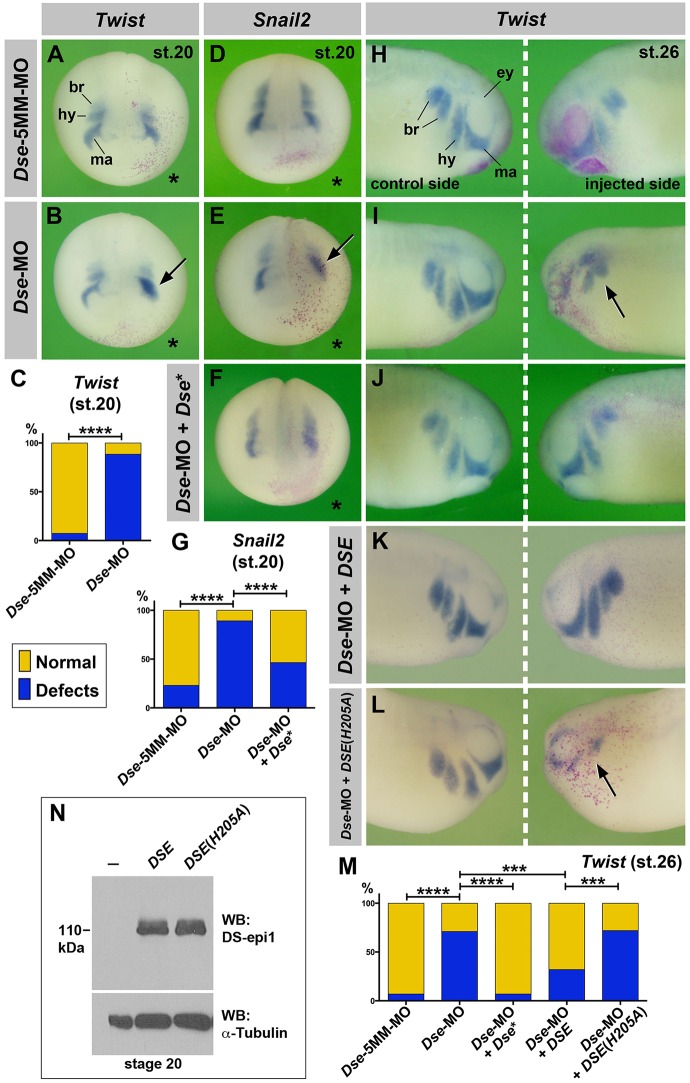
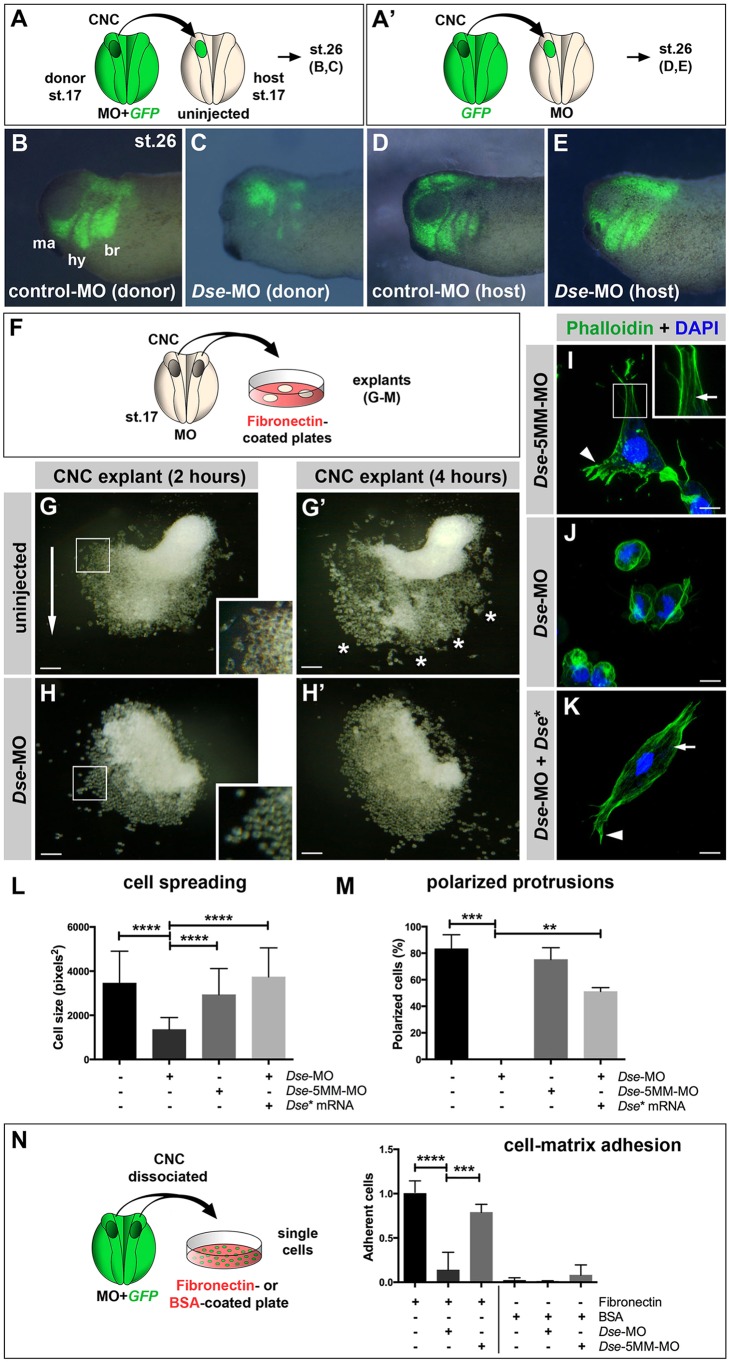
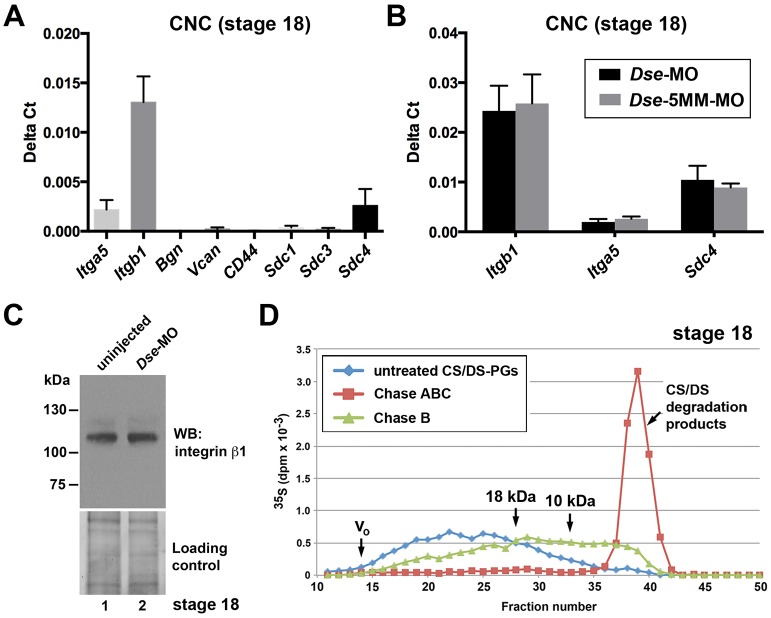
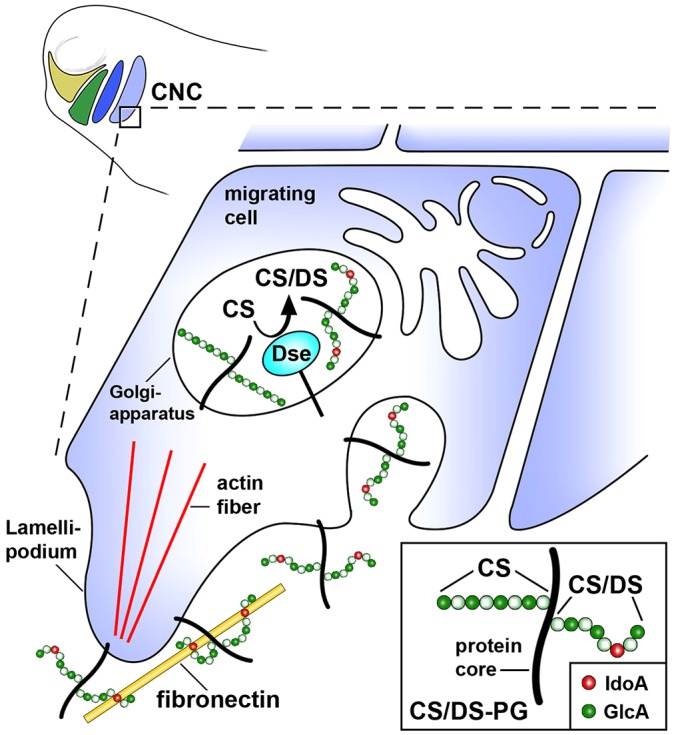
Of all live births with congenital anomalies, approximately one-third exhibit deformities of the head and face. Most craniofacial disorders are associated with defects in a migratory stem and progenitor cell population, which is designated the neural crest (NC). Musculocontractural Ehlers-Danlos syndrome (MCEDS) is a heritable connective tissue disorder with distinct craniofacial features; this syndrome comprises multiple congenital malformations that are caused by dysfunction of dermatan sulfate (DS) biosynthetic enzymes, including DS epimerase-1 (DS-epi1; also known as DSE). Studies in mice have extended our understanding of DS-epi1 in connective tissue maintenance; however, its role in fetal development is not understood. We demonstrate that DS-epi1 is important for the generation of isolated iduronic acid residues in chondroitin sulfate (CS)/DS proteoglycans in early Xenopus embryos. The knockdown of DS-epi1 does not affect the formation of early NC progenitors; however, it impairs the correct activation of transcription factors involved in the epithelial-mesenchymal transition (EMT) and reduces the extent of NC cell migration, which leads to a decrease in NC-derived craniofacial skeleton, melanocytes and dorsal fin structures. Transplantation experiments demonstrate a tissue-autonomous role for DS-epi1 in cranial NC cell migration in vivo Cranial NC explant and single-cell cultures indicate a requirement of DS-epi1 in cell adhesion, spreading and extension of polarized cell processes on fibronectin. Thus, our work indicates a functional link between DS and NC cell migration. We conclude that NC defects in the EMT and cell migration might account for the craniofacial anomalies and other congenital malformations in MCEDS, which might facilitate the diagnosis and development of therapies for this distressing condition. Moreover, the presented correlations between human DS-epi1 expression and gene sets of mesenchymal character, invasion and metastasis in neuroblastoma and malignant melanoma suggest an association between DS and NC-derived cancers.
Keywords: Cancer; Cell migration; Dermatan sulfate; Musculocontractural Ehlers–Danlos syndrome; Neural crest; Xenopus.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Bartolini B., Thelin M. A., Rauch U., Feinstein R., Oldberg Å., Malmström A. and Maccarana M. (2012). Mouse development is not obviously affected by the absence of dermatan sulfate epimerase 2 in spite of a modified brain dermatan sulfate composition. Glycobiology 22, 1007-1016. 10.1093/glycob/cws065 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
