

Detyrosinated microtubules buckle and bear load in contracting cardiomyocytes
- PMID: 27102488
- PMCID: PMC5441927
- DOI: 10.1126/science.aaf0659
Detyrosinated microtubules buckle and bear load in contracting cardiomyocytes
Abstract
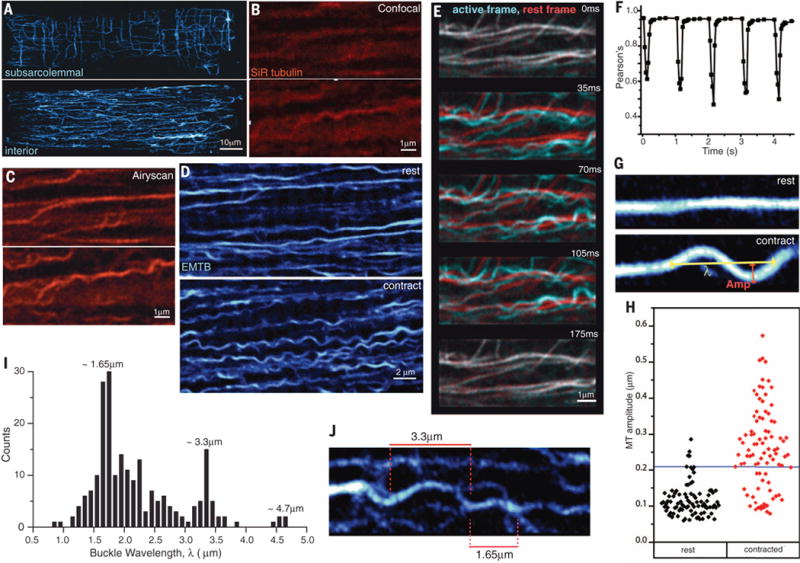
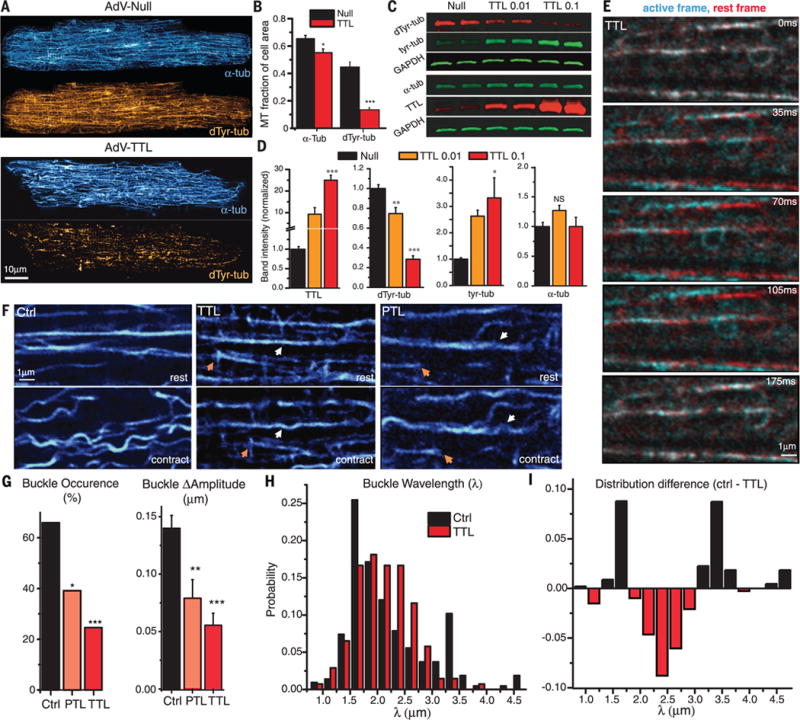
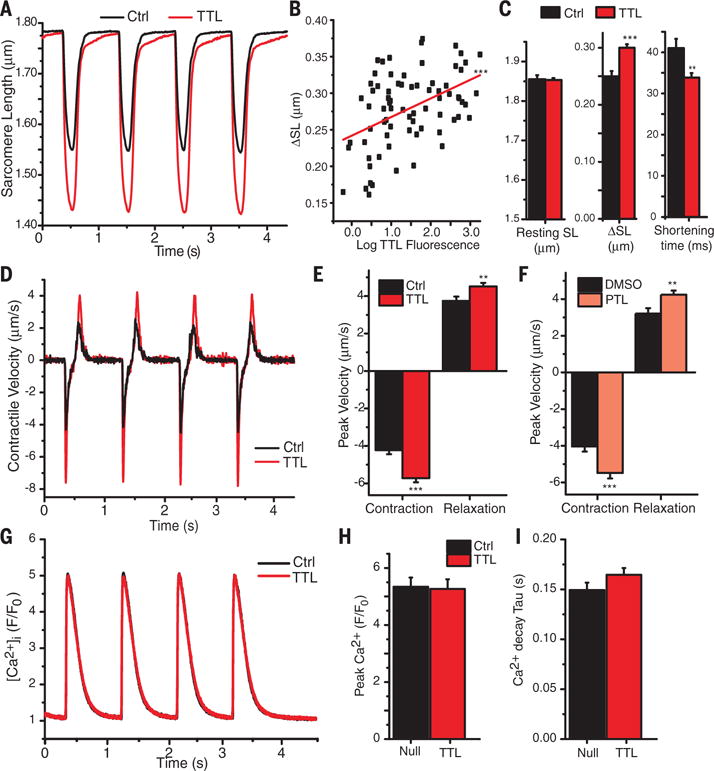
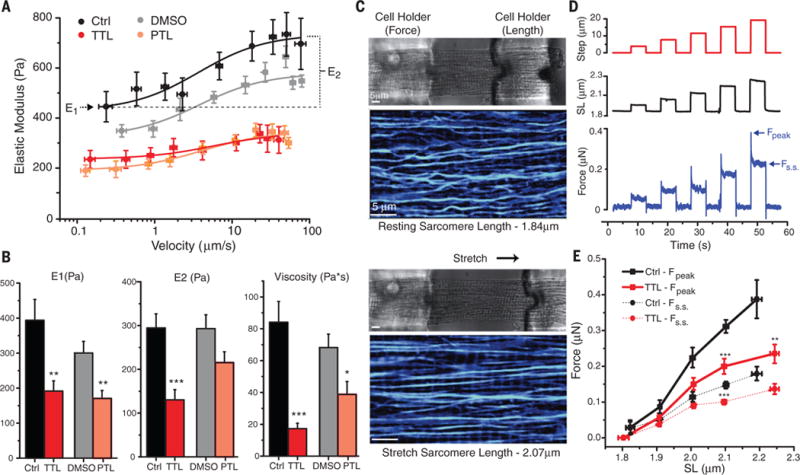
The microtubule (MT) cytoskeleton can transmit mechanical signals and resist compression in contracting cardiomyocytes. How MTs perform these roles remains unclear because of difficulties in observing MTs during the rapid contractile cycle. Here, we used high spatial and temporal resolution imaging to characterize MT behavior in beating mouse myocytes. MTs deformed under contractile load into sinusoidal buckles, a behavior dependent on posttranslational "detyrosination" of α-tubulin. Detyrosinated MTs associated with desmin at force-generating sarcomeres. When detyrosination was reduced, MTs uncoupled from sarcomeres and buckled less during contraction, which allowed sarcomeres to shorten and stretch with less resistance. Conversely, increased detyrosination promoted MT buckling, stiffened the myocyte, and correlated with impaired function in cardiomyopathy. Thus, detyrosinated MTs represent tunable, compression-resistant elements that may impair cardiac function in disease.
Copyright © 2016, American Association for the Advancement of Science.
Figures
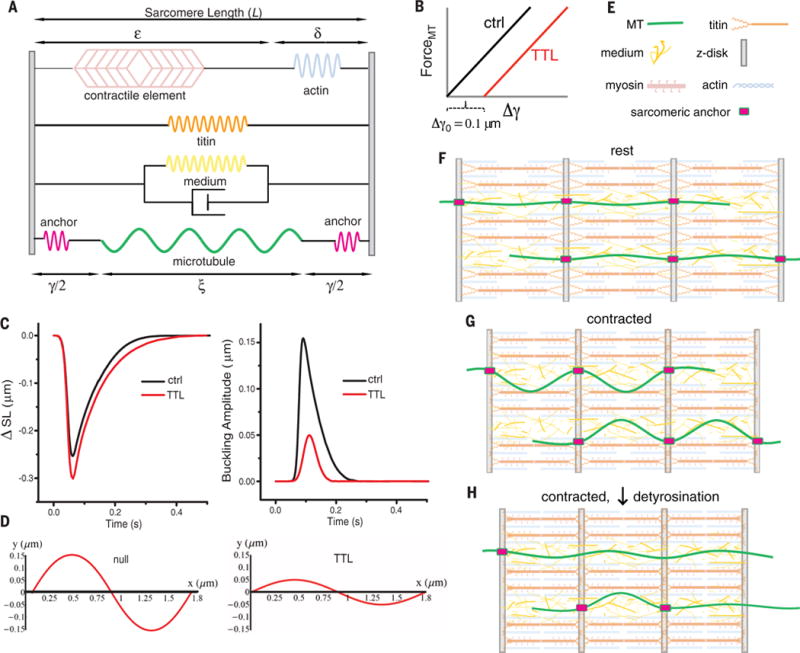
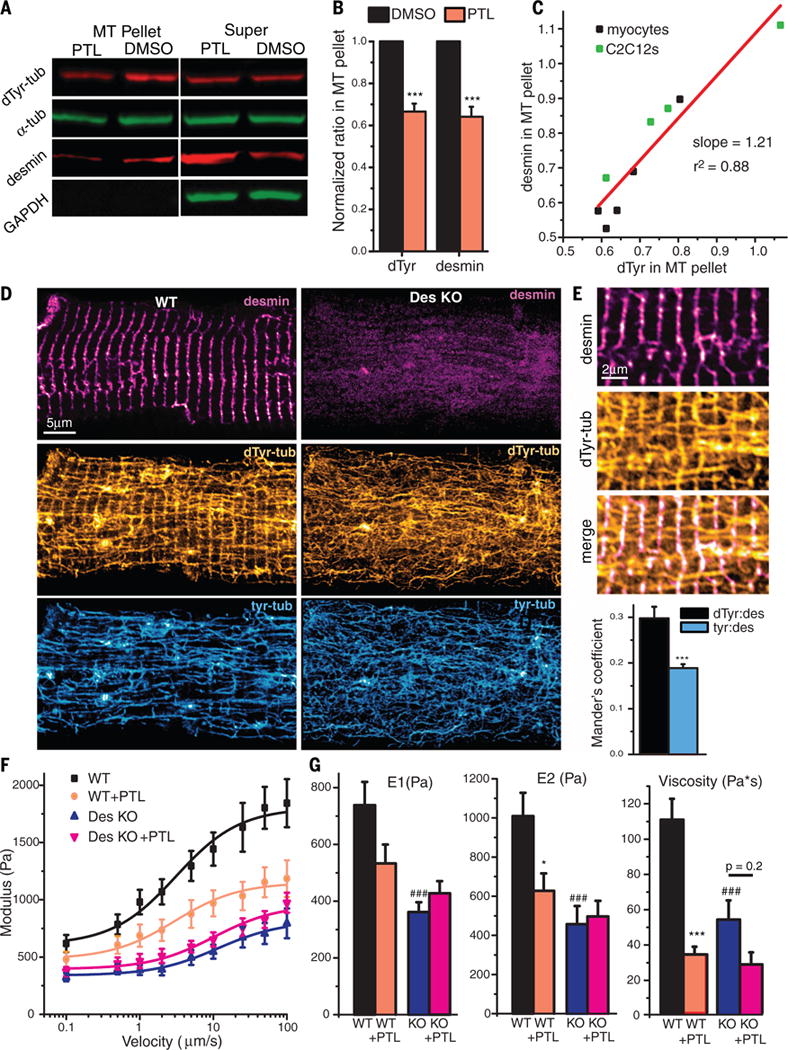
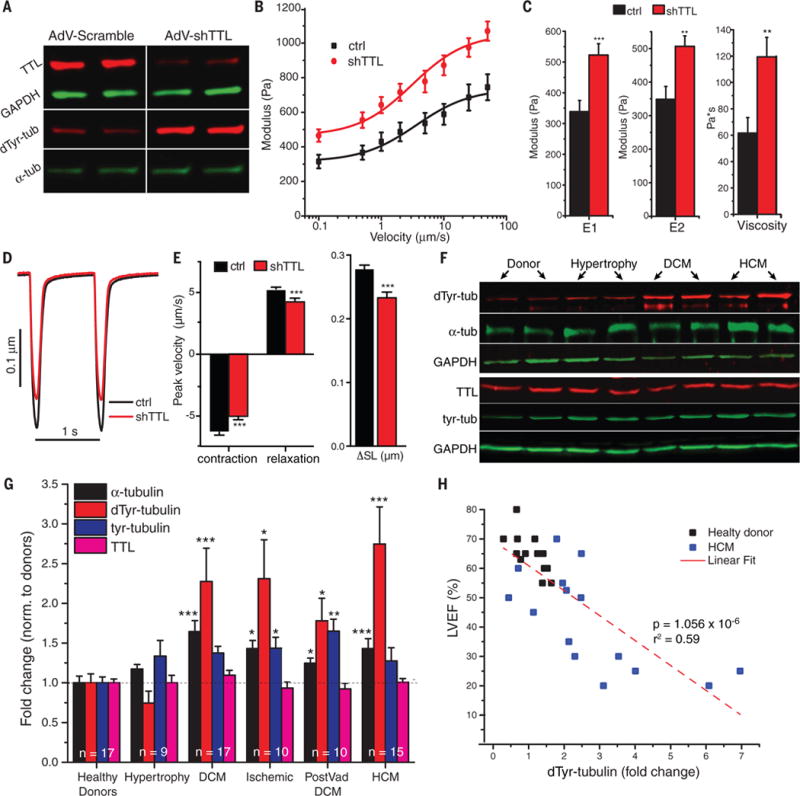
Comment in
-
Cytoskeleton: Microtubules set the beat.Nat Rev Mol Cell Biol. 2016 Jun;17(6):333. doi: 10.1038/nrm.2016.63. Epub 2016 May 11. Nat Rev Mol Cell Biol. 2016. PMID: 27165791 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
- HL089847/HL/NHLBI NIH HHS/United States
- HL105993/HL/NHLBI NIH HHS/United States
- R00-HL114879/HL/NHLBI NIH HHS/United States
- T32HL007954/HL/NHLBI NIH HHS/United States
- R01 HL105993/HL/NHLBI NIH HHS/United States
- T32AR053461-09/AR/NIAMS NIH HHS/United States
- T32 AR053461/AR/NIAMS NIH HHS/United States
- R01 EB017753/EB/NIBIB NIH HHS/United States
- R00 HL114879/HL/NHLBI NIH HHS/United States
- R01 HL089847/HL/NHLBI NIH HHS/United States
- R01EB017753/EB/NIBIB NIH HHS/United States
- R01 HL133080/HL/NHLBI NIH HHS/United States
- T32 HL007954/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
