

Hybrid Sterility Locus on Chromosome X Controls Meiotic Recombination Rate in Mouse
- PMID: 27104744
- PMCID: PMC4841592
- DOI: 10.1371/journal.pgen.1005906
Hybrid Sterility Locus on Chromosome X Controls Meiotic Recombination Rate in Mouse
Abstract
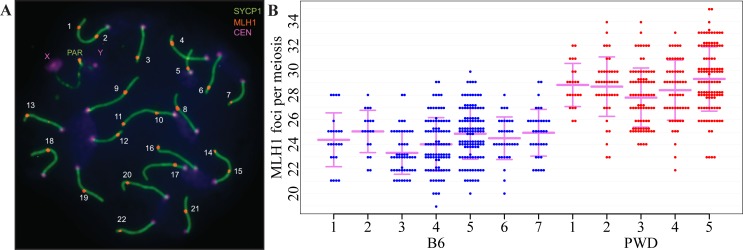
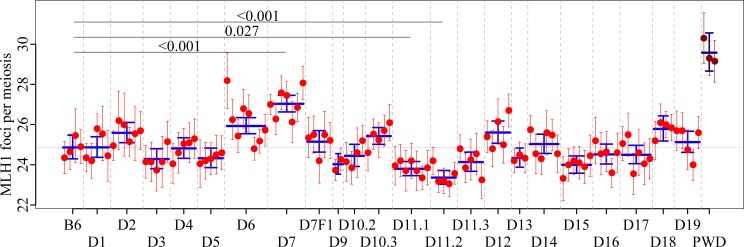
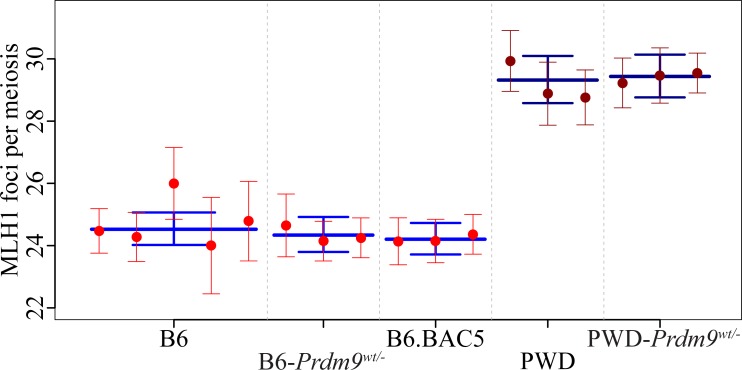
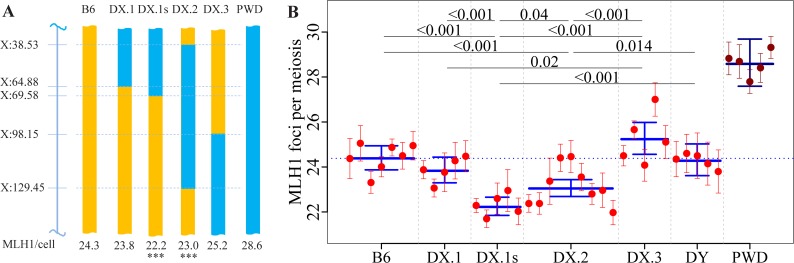
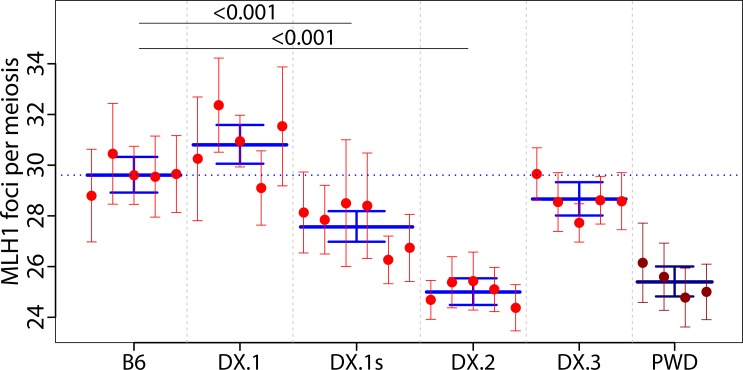
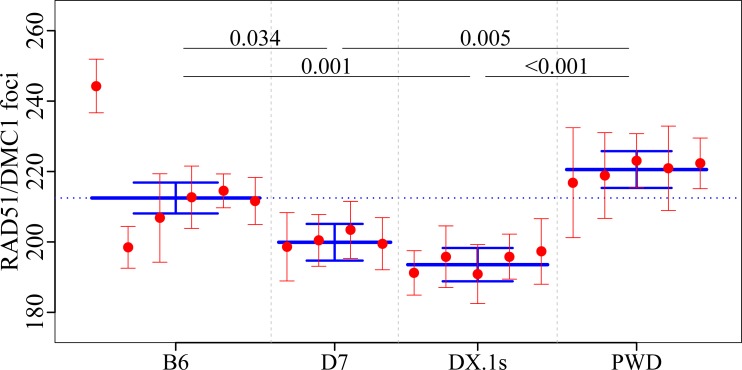
Meiotic recombination safeguards proper segregation of homologous chromosomes into gametes, affects genetic variation within species, and contributes to meiotic chromosome recognition, pairing and synapsis. The Prdm9 gene has a dual role, it controls meiotic recombination by determining the genomic position of crossover hotspots and, in infertile hybrids of house mouse subspecies Mus m. musculus (Mmm) and Mus m. domesticus (Mmd), it further functions as the major hybrid sterility gene. In the latter role Prdm9 interacts with the hybrid sterility X 2 (Hstx2) genomic locus on Chromosome X (Chr X) by a still unknown mechanism. Here we investigated the meiotic recombination rate at the genome-wide level and its possible relation to hybrid sterility. Using immunofluorescence microscopy we quantified the foci of MLH1 DNA mismatch repair protein, the cytological counterparts of reciprocal crossovers, in a panel of inter-subspecific chromosome substitution strains. Two autosomes, Chr 7 and Chr 11, significantly modified the meiotic recombination rate, yet the strongest modifier, designated meiotic recombination 1, Meir1, emerged in the 4.7 Mb Hstx2 genomic locus on Chr X. The male-limited transgressive effect of Meir1 on recombination rate parallels the male-limited transgressive role of Hstx2 in hybrid male sterility. Thus, both genetic factors, the Prdm9 gene and the Hstx2/Meir1 genomic locus, indicate a link between meiotic recombination and hybrid sterility. A strong female-specific modifier of meiotic recombination rate with the effect opposite to Meir1 was localized on Chr X, distally to Meir1. Mapping Meir1 to a narrow candidate interval on Chr X is an important first step towards positional cloning of the respective gene(s) responsible for variation in the global recombination rate between closely related mouse subspecies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Moens PB, Kolas NK, Tarsounas M, Marcon E, Cohen PE, et al. (2002) The time course and chromosomal localization of recombination-related proteins at meiosis in the mouse are compatible with models that can resolve the early DNA-DNA interactions without reciprocal recombination. J Cell Sci 115: 1611–1622. - PubMed
-
- Bellani MA, Boateng KA, McLeod D, Camerini-Otero RD (2010) The expression profile of the major mouse SPO11 isoforms indicates that SPO11beta introduces double strand breaks and suggests that SPO11alpha has an additional role in prophase in both spermatocytes and oocytes. Mol Cell Biol 30: 4391–4403. 10.1128/MCB.00002-10 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
