

Functional Implications of Human-Specific Changes in Great Ape microRNAs
- PMID: 27105073
- PMCID: PMC4841587
- DOI: 10.1371/journal.pone.0154194
Functional Implications of Human-Specific Changes in Great Ape microRNAs
Abstract
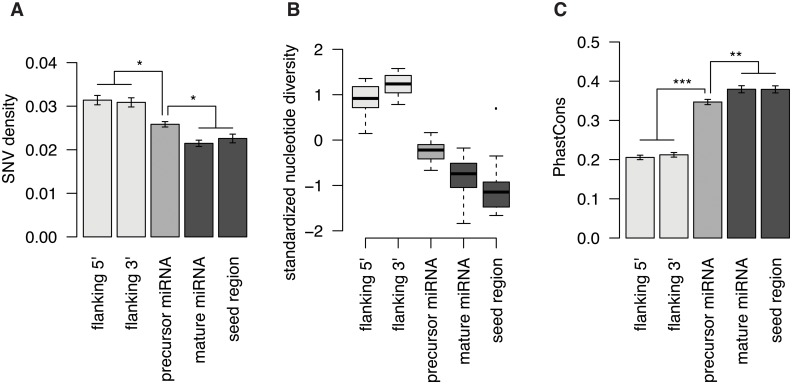
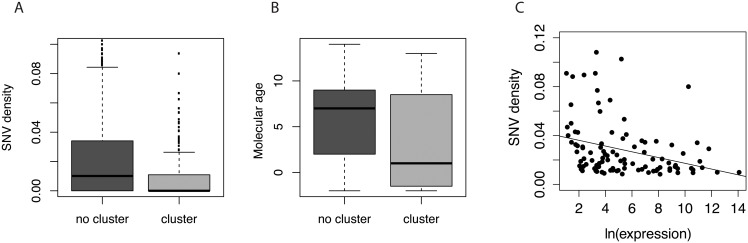
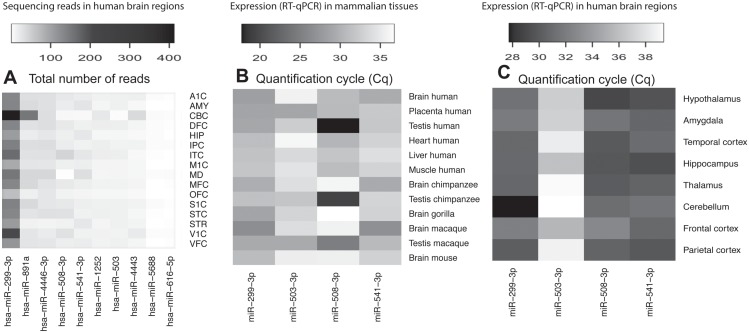
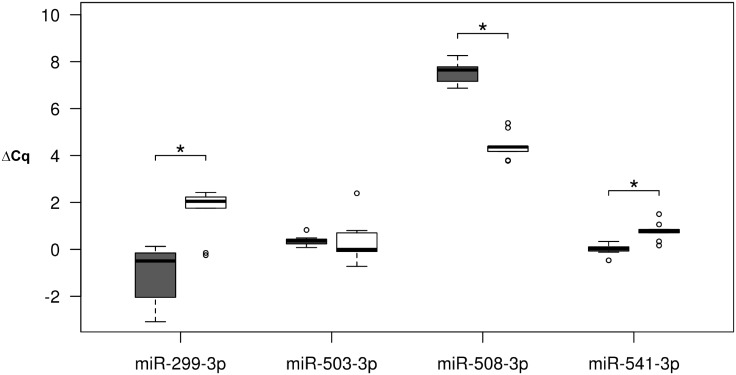
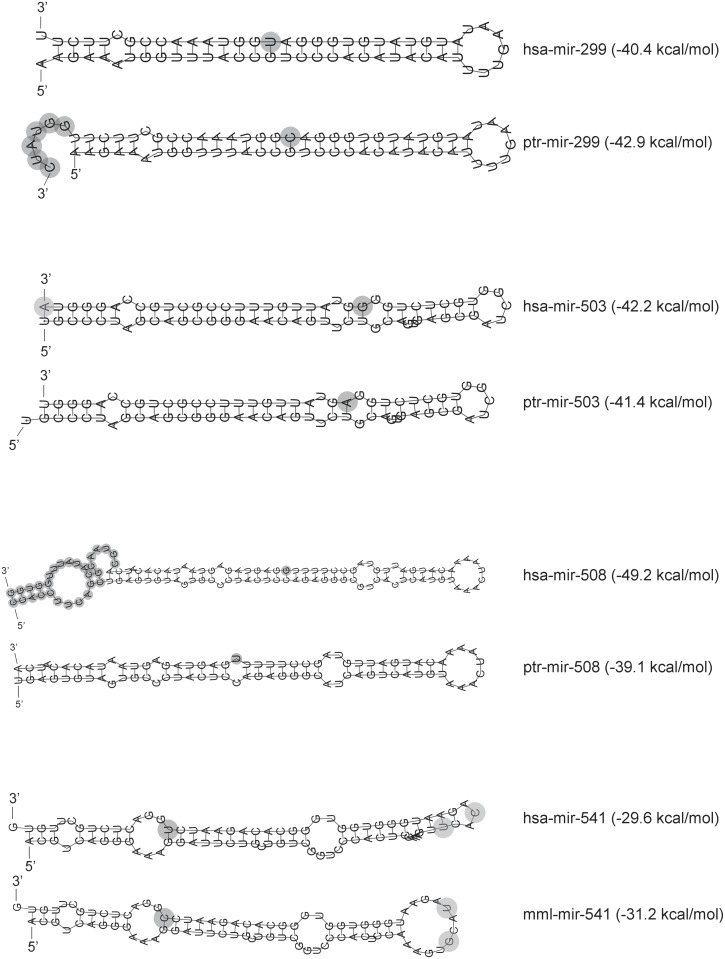
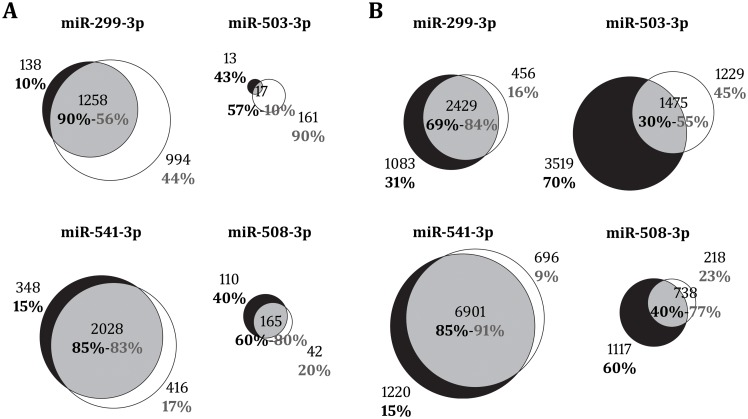
microRNAs are crucial post-transcriptional regulators of gene expression involved in a wide range of biological processes. Although microRNAs are highly conserved among species, the functional implications of existing lineage-specific changes and their role in determining differences between humans and other great apes have not been specifically addressed. We analyzed the recent evolutionary history of 1,595 human microRNAs by looking at their intra- and inter-species variation in great apes using high-coverage sequenced genomes of 82 individuals including gorillas, orangutans, bonobos, chimpanzees and humans. We explored the strength of purifying selection among microRNA regions and found that the seed and mature regions are under similar and stronger constraint than the precursor region. We further constructed a comprehensive catalogue of microRNA species-specific nucleotide substitutions among great apes and, for the first time, investigated the biological relevance that human-specific changes in microRNAs may have had in great ape evolution. Expression and functional analyses of four microRNAs (miR-299-3p, miR-503-3p, miR-508-3p and miR-541-3p) revealed that lineage-specific nucleotide substitutions and changes in the length of these microRNAs alter their expression as well as the repertoires of target genes and regulatory networks. We suggest that the studied molecular changes could have modified crucial microRNA functions shaping phenotypes that, ultimately, became human-specific. Our work provides a frame to study the impact that regulatory changes may have in the recent evolution of our species.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
