

Reconstruction of Cell Surface Densities of Ion Pumps, Exchangers, and Channels from mRNA Expression, Conductance Kinetics, Whole-Cell Calcium, and Current-Clamp Voltage Recordings, with an Application to Human Uterine Smooth Muscle Cells
- PMID: 27105427
- PMCID: PMC4841602
- DOI: 10.1371/journal.pcbi.1004828
Reconstruction of Cell Surface Densities of Ion Pumps, Exchangers, and Channels from mRNA Expression, Conductance Kinetics, Whole-Cell Calcium, and Current-Clamp Voltage Recordings, with an Application to Human Uterine Smooth Muscle Cells
Abstract
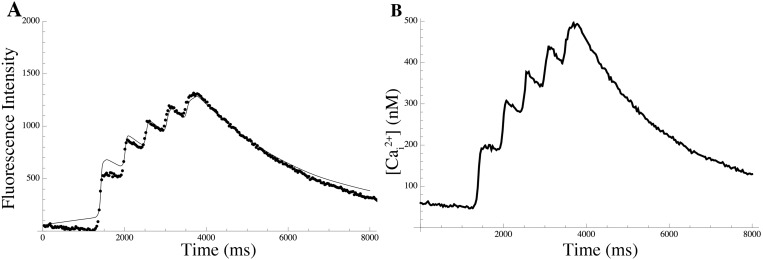
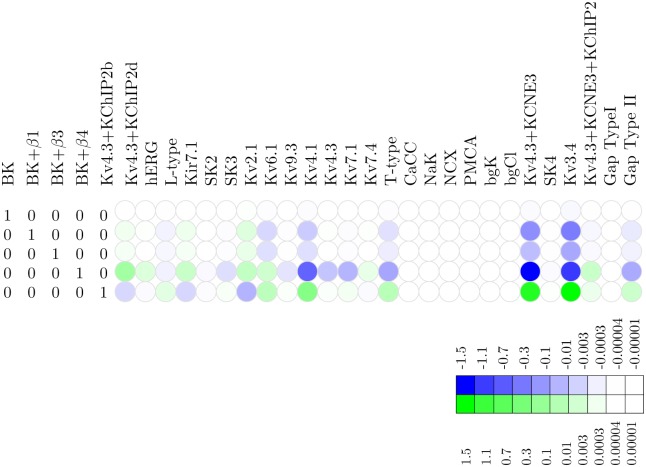
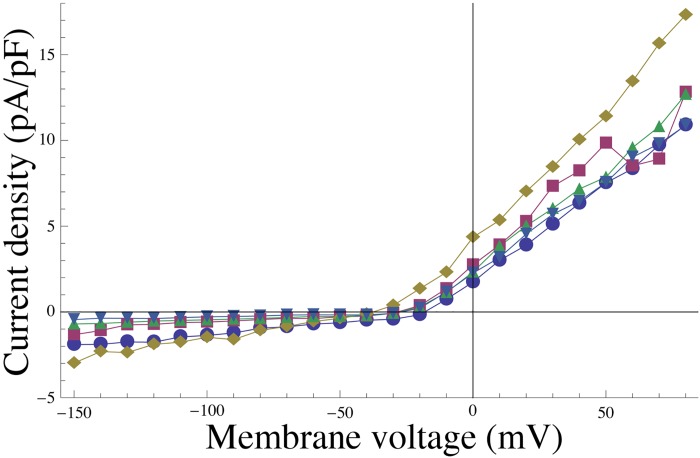
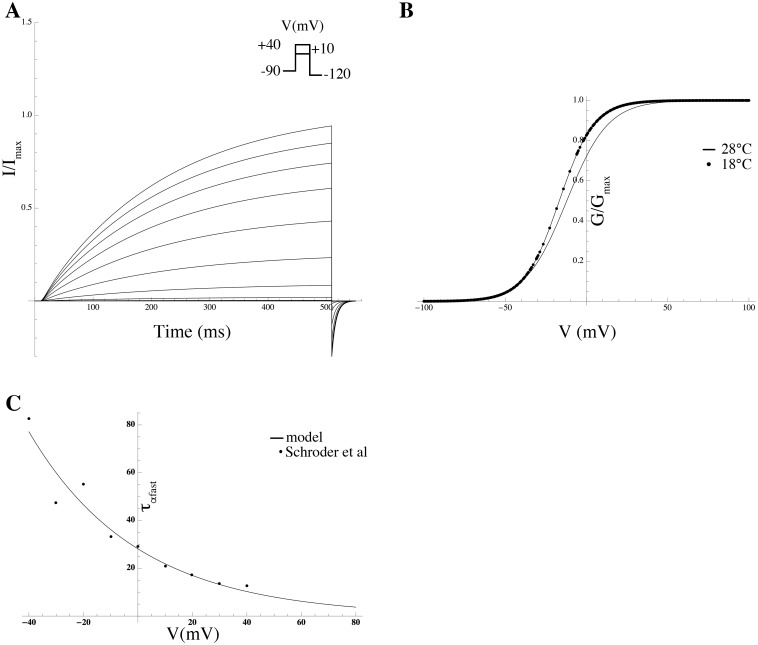
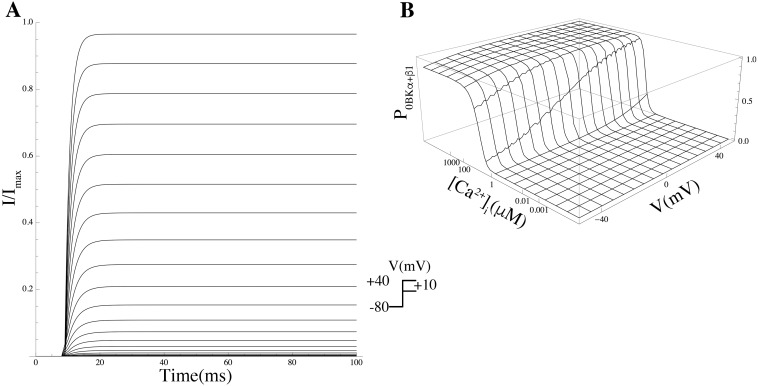
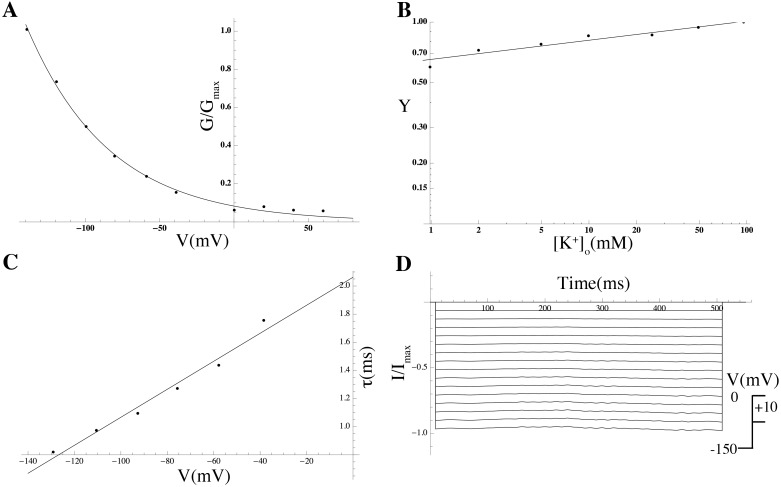
Uterine smooth muscle cells remain quiescent throughout most of gestation, only generating spontaneous action potentials immediately prior to, and during, labor. This study presents a method that combines transcriptomics with biophysical recordings to characterise the conductance repertoire of these cells, the 'conductance repertoire' being the total complement of ion channels and transporters expressed by an electrically active cell. Transcriptomic analysis provides a set of potential electrogenic entities, of which the conductance repertoire is a subset. Each entity within the conductance repertoire was modeled independently and its gating parameter values were fixed using the available biophysical data. The only remaining free parameters were the surface densities for each entity. We characterise the space of combinations of surface densities (density vectors) consistent with experimentally observed membrane potential and calcium waveforms. This yields insights on the functional redundancy of the system as well as its behavioral versatility. Our approach couples high-throughput transcriptomic data with physiological behaviors in health and disease, and provides a formal method to link genotype to phenotype in excitable systems. We accurately predict current densities and chart functional redundancy. For example, we find that to evoke the observed voltage waveform, the BK channel is functionally redundant whereas hERG is essential. Furthermore, our analysis suggests that activation of calcium-activated chloride conductances by intracellular calcium release is the key factor underlying spontaneous depolarisations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





































References
-
- Hille B (2001) Ion Channels of Excitable Membranes, Third Edition Sinauer Associates, Inc.
-
- ten Tusscher K, Noble D, Noble PJ, Panfilov AV (2004) A model for human ventricular tissue. Am J Physiol Heart Circ Physiol 286: H1573–H1589. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
