

Strategies of Mesenchymal Invasion of Patient-derived Brain Tumors: Microenvironmental Adaptation
- PMID: 27108713
- PMCID: PMC4842976
- DOI: 10.1038/srep24912
Strategies of Mesenchymal Invasion of Patient-derived Brain Tumors: Microenvironmental Adaptation
Abstract
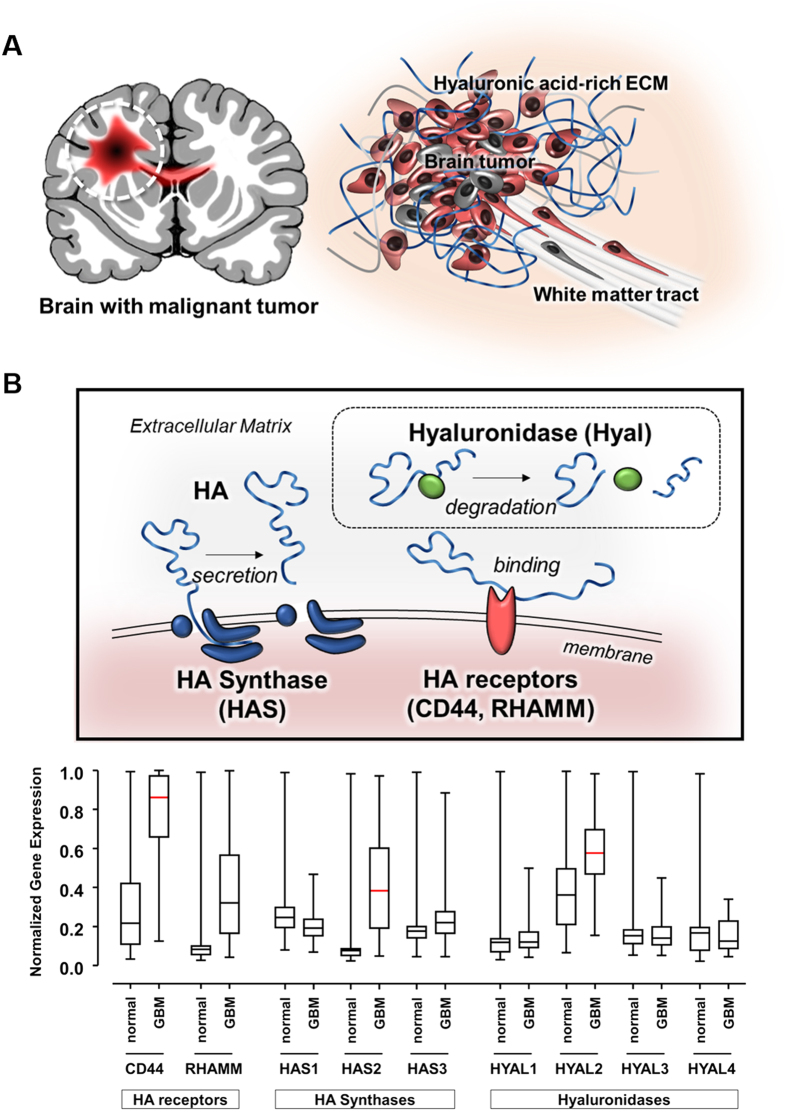
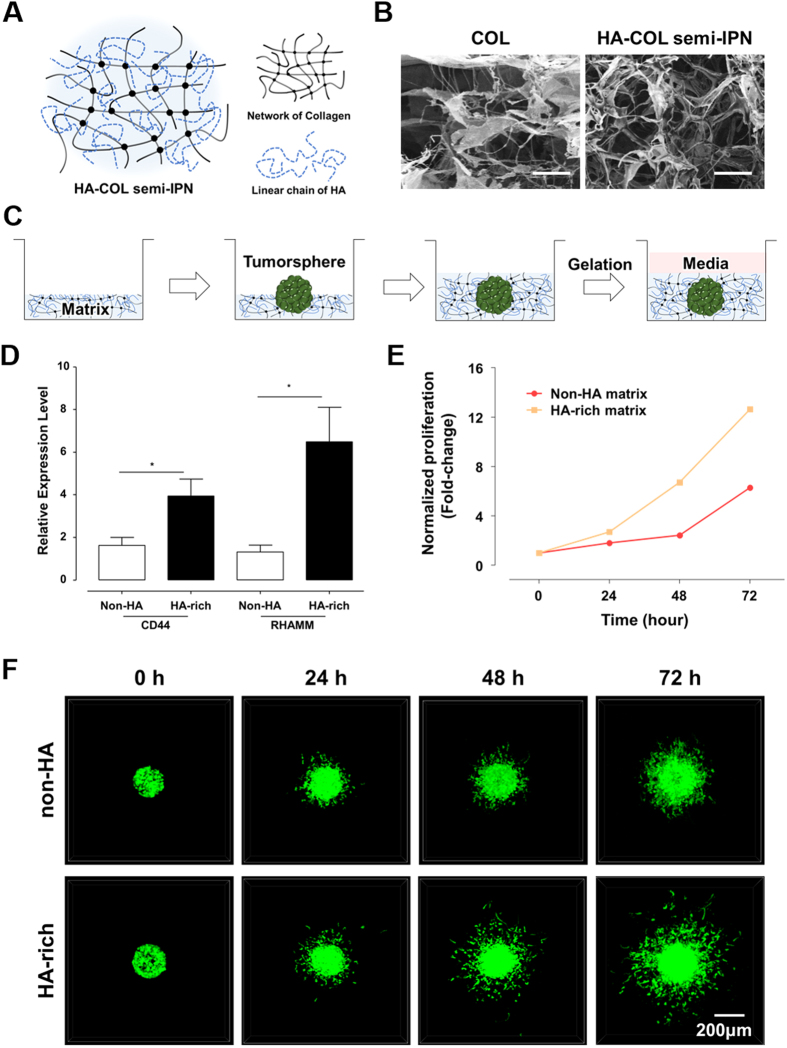
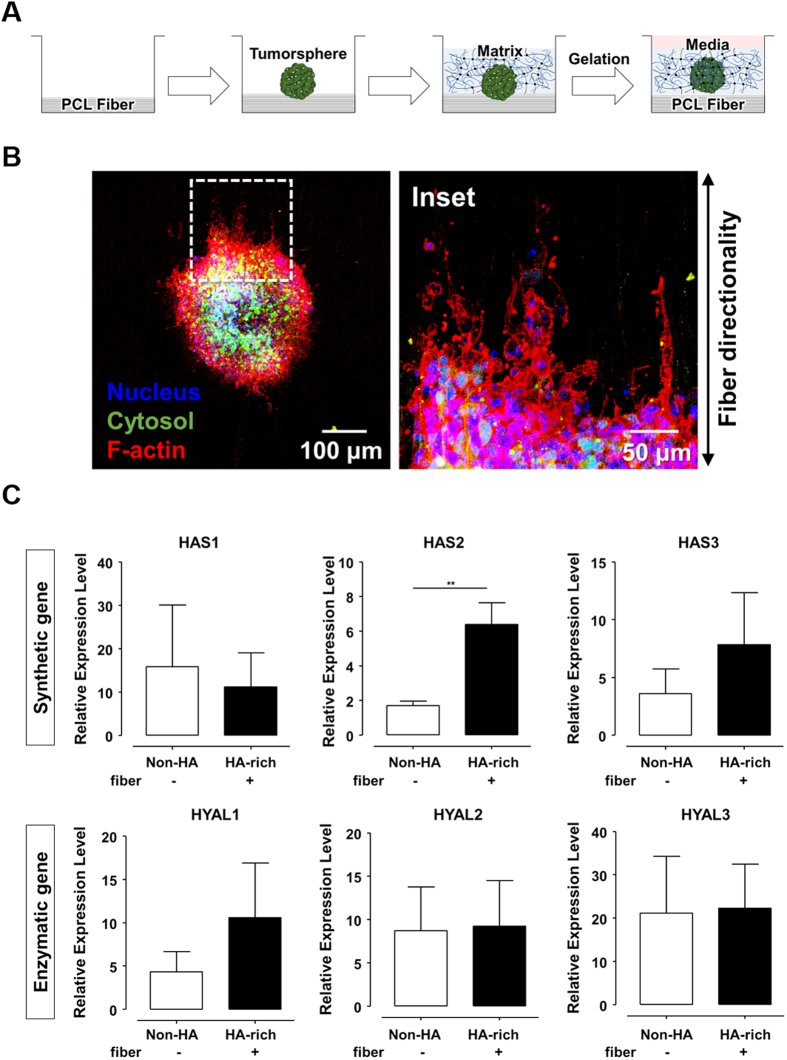
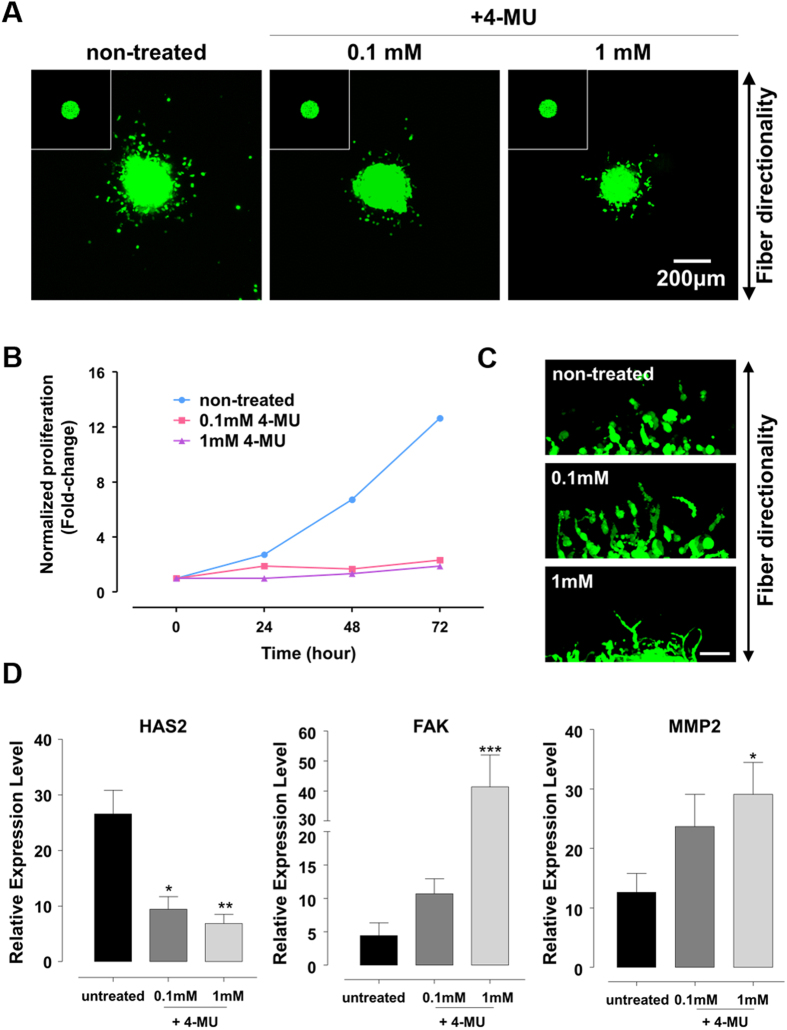
The high mortality in glioblastoma multiforme (GBM) patients is primarily caused by extensive infiltration into adjacent tissue and subsequent rapid recurrence. There are no clear therapeutic strategies that target the infiltrative subpopulation of GBM mass. Using mesenchymal mode of invasion, the GBM is known to widely infiltrate by interacting with various unique components within brain microenvironment such as hyaluronic acid (HA)-rich matrix and white matter tracts. However, it is unclear how these GBM microenvironments influence the strategies of mesenchymal invasion. We hypothesize that GBM has different strategies to facilitate such invasion through adaptation to their local microenvironment. Using our in vitro biomimetic microenvironment platform for three-dimensional GBM tumorspheres (TSs), we found that the strategies of GBM invasion were predominantly regulated by the HA-rich ECM microenvironment, showing marked phenotypic changes in the presence of HA, which were mainly mediated by HA synthase (HAS). Interestingly, after inhibition of the HAS gene, GBM switched their invasion strategies to a focal adhesion (FA)-mediated invasion. These results demonstrate that the microenvironmental adaptation allowed a flexible invasion strategy for GBM. Using our model, we suggest a new inhibitory pathway for targeting infiltrative GBM and propose an importance of multi-target therapy for GBM, which underwent microenvironmental adaptation.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
