

Concerted genomic targeting of H3K27 demethylase REF6 and chromatin-remodeling ATPase BRM in Arabidopsis
- PMID: 27111034
- PMCID: PMC5134324
- DOI: 10.1038/ng.3555
Concerted genomic targeting of H3K27 demethylase REF6 and chromatin-remodeling ATPase BRM in Arabidopsis
Abstract
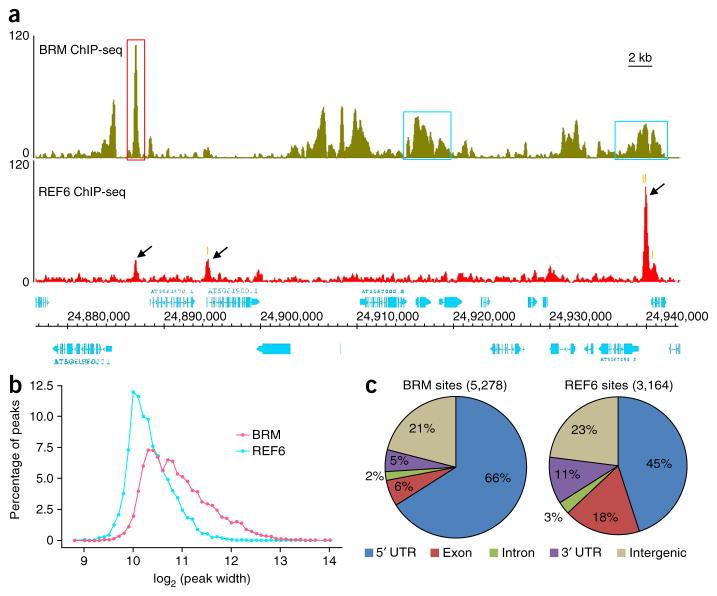
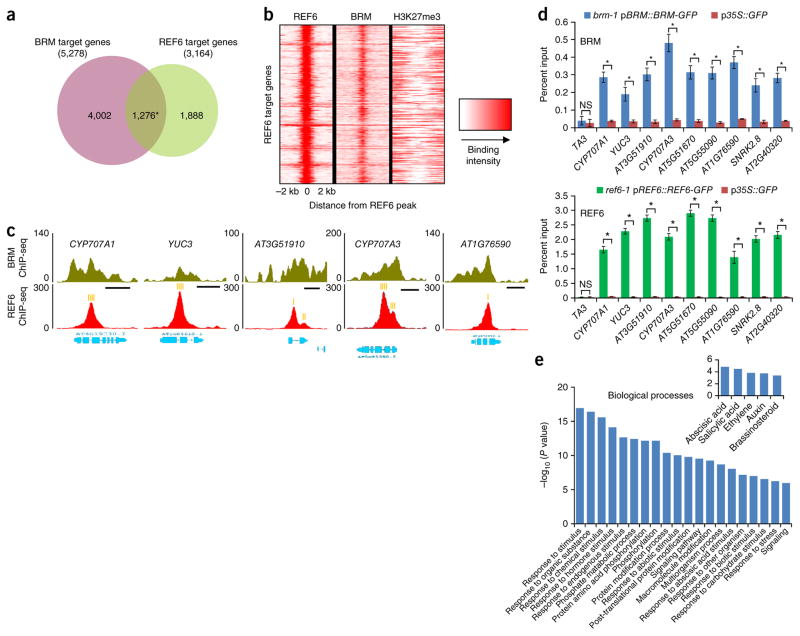
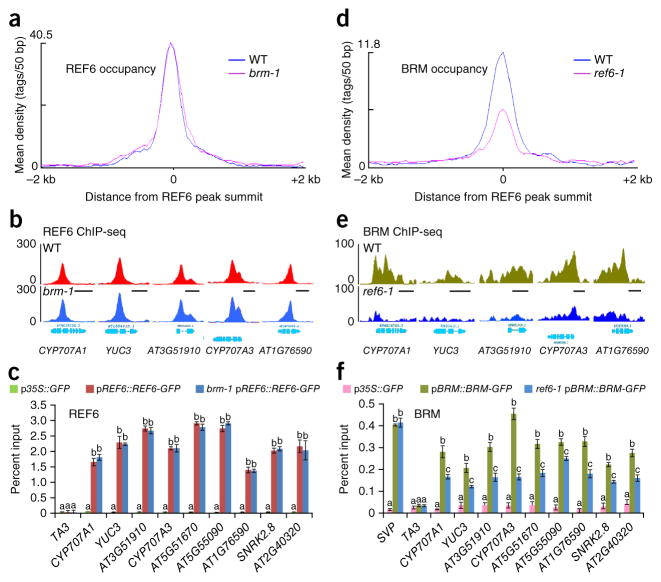
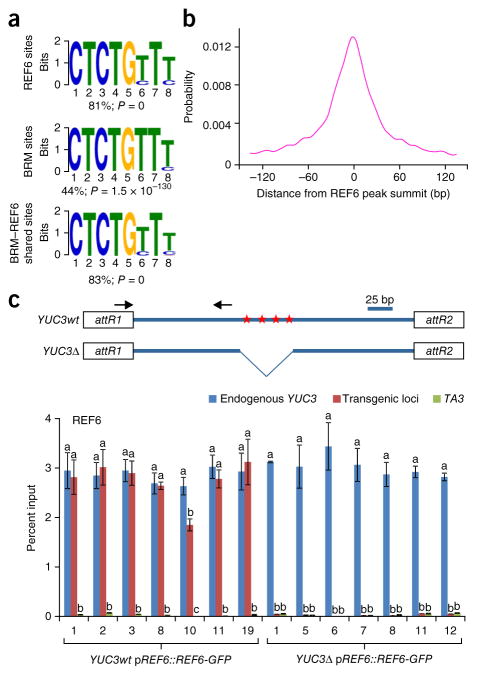
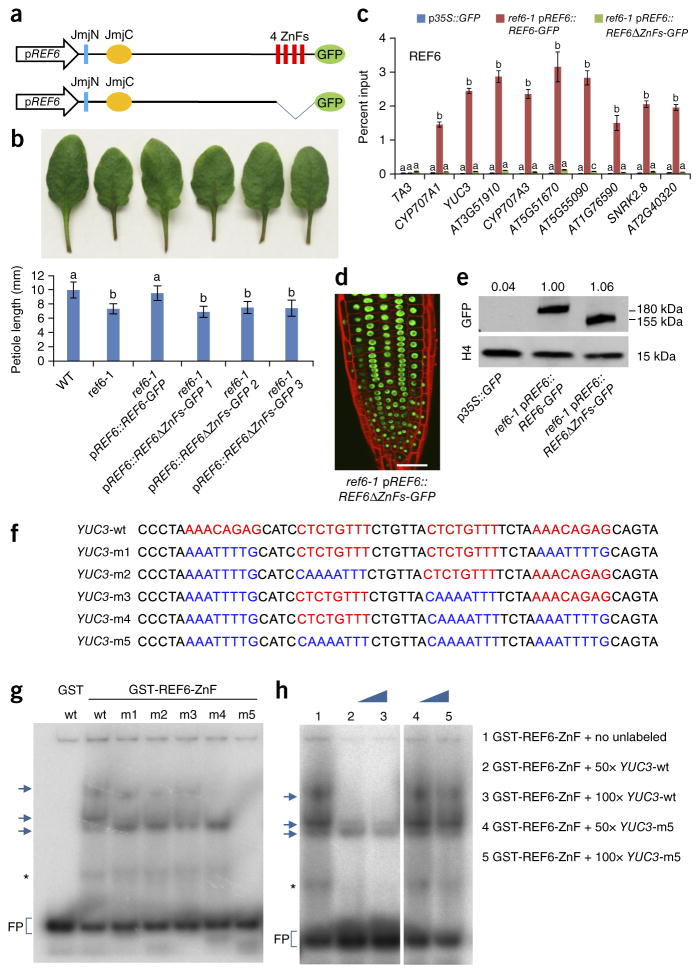
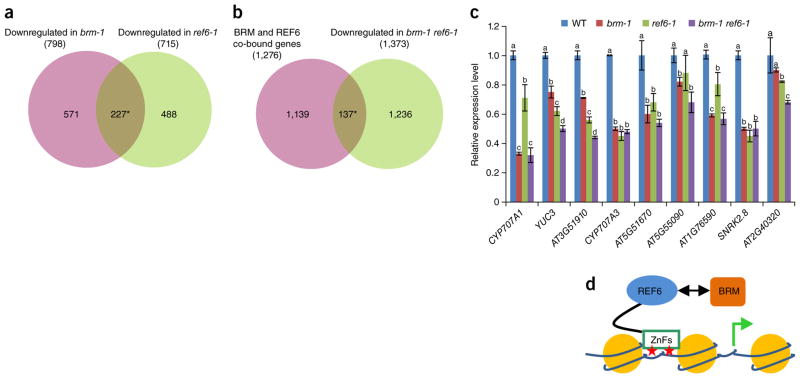
SWI/SNF-type chromatin remodelers, such as BRAHMA (BRM), and H3K27 demethylases both have active roles in regulating gene expression at the chromatin level, but how they are recruited to specific genomic sites remains largely unknown. Here we show that RELATIVE OF EARLY FLOWERING 6 (REF6), a plant-unique H3K27 demethylase, targets genomic loci containing a CTCTGYTY motif via its zinc-finger (ZnF) domains and facilitates the recruitment of BRM. Genome-wide analyses showed that REF6 colocalizes with BRM at many genomic sites with the CTCTGYTY motif. Loss of REF6 results in decreased BRM occupancy at BRM-REF6 co-targets. Furthermore, REF6 directly binds to the CTCTGYTY motif in vitro, and deletion of the motif from a target gene renders it inaccessible to REF6 in vivo. Finally, we show that, when its ZnF domains are deleted, REF6 loses its genomic targeting ability. Thus, our work identifies a new genomic targeting mechanism for an H3K27 demethylase and demonstrates its key role in recruiting the BRM chromatin remodeler.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
DNA-sequence-specific erasers of epigenetic memory.Nat Genet. 2016 May 27;48(6):591-2. doi: 10.1038/ng.3579. Nat Genet. 2016. PMID: 27230685
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
