

Complex Genetics of Type 2 Diabetes and Effect Size: What have We Learned from Isolated Populations?
- PMID: 27111117
- PMCID: PMC5275756
- DOI: 10.1900/RDS.2015.12.299
Complex Genetics of Type 2 Diabetes and Effect Size: What have We Learned from Isolated Populations?
Abstract
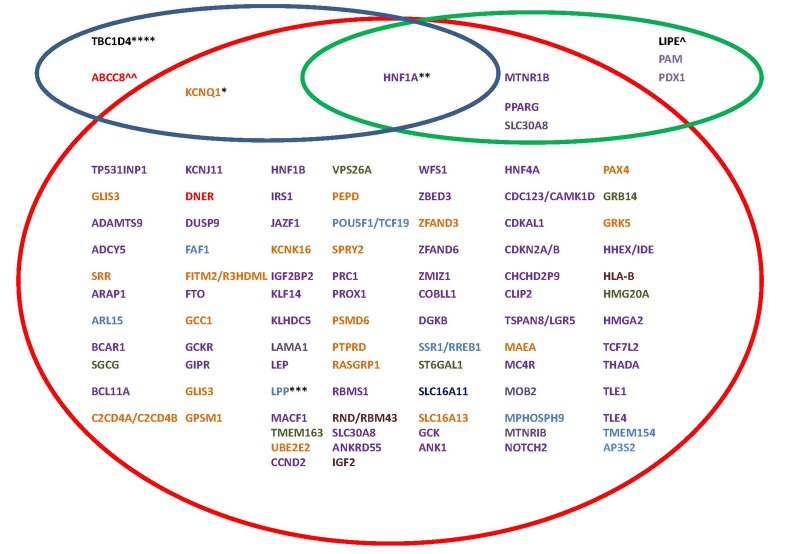
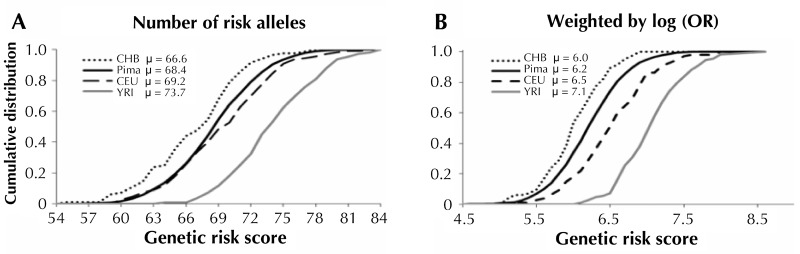
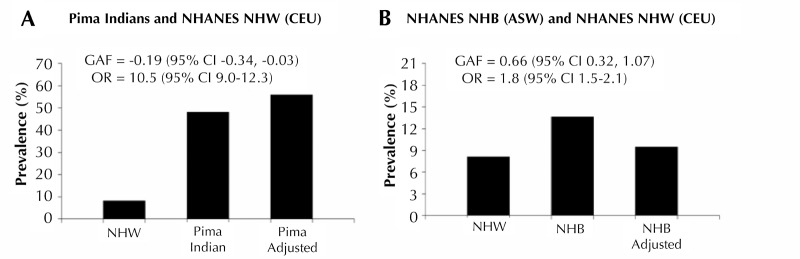
Genetic studies in large outbred populations have documented a complex, highly polygenic basis for type 2 diabetes (T2D). Most of the variants currently known to be associated with T2D risk have been identified in large studies that included tens of thousands of individuals who are representative of a single major ethnic group such as European, Asian, or African. However, most of these variants have only modest effects on the risk for T2D; identification of definitive 'causal variant' or 'causative loci' is typically lacking. Studies in isolated populations offer several advantages over outbred populations despite being, on average, much smaller in sample size. For example, reduced genetic variability, enrichment of rare variants, and a more uniform environment and lifestyle, which are hallmarks of isolated populations, can reduce the complexity of identifying disease-associated genes. To date, studies in isolated populations have provided valuable insight into the genetic basis of T2D by providing both a deeper understanding of previously identified T2D-associated variants (e.g. demonstrating that variants in KCNQ1 have a strong parent-of-origin effect) or providing novel variants (e.g. ABCC8 in Pima Indians, TBC1D4 in the Greenlandic population, HNF1A in Canadian Oji-Cree). This review summarizes advancements in genetic studies of T2D in outbred and isolated populations, and provides information on whether the difference in the prevalence of T2D in different populations (Pima Indians vs. non-Hispanic Whites and non-Hispanic Whites vs. non-Hispanic Blacks) can be explained by the difference in risk allele frequencies of established T2D variants.
Conflict of interest statement
The authors report no conflict of interests.
Figures
References
-
- Lillioja S, Mott DM, Spraul M, Ferraro R, Foley JE, Ravussin E, Knowler WC, Bennett PH, Bogardus C. Insulin resistance and insulin secretory dysfunction as precursors of non-insulin-dependent diabetes mellitus. Prospective studies of Pima Indians. N Engl J Med. 1993;329(27):1988–1992. - PubMed
-
- Mayer-Davis EJ, Costacou T. Obesity and sedentary lifestyle: modifiable risk factors for prevention of type 2 diabetes. Curr Diab Rep. 2001;1(2):170–176. - PubMed
-
- Meigs JB, Cupples LA, Wilson PW. Parental transmission of type 2 diabetes: the Framingham Offspring Study. Diabetes. 2000;49(12):2201–2207. - PubMed
-
- Barnett AH, Eff C, Leslie RD, Pyke DA. Diabetes in identical twins. A study of 200 pairs. Diabetologia. 1981;20(2):87–93. - PubMed
-
- Newman B, Selby JV, King MC, Slemenda C, Fabsitz R, Friedman GD. Concordance for type 2 (non-insulin-dependent) diabetes mellitus in male twins. Diabetologia. 1987;30(10):763–768. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical