

The Limited Utility of Multiunit Data in Differentiating Neuronal Population Activity
- PMID: 27111446
- PMCID: PMC4844128
- DOI: 10.1371/journal.pone.0153154
The Limited Utility of Multiunit Data in Differentiating Neuronal Population Activity
Abstract
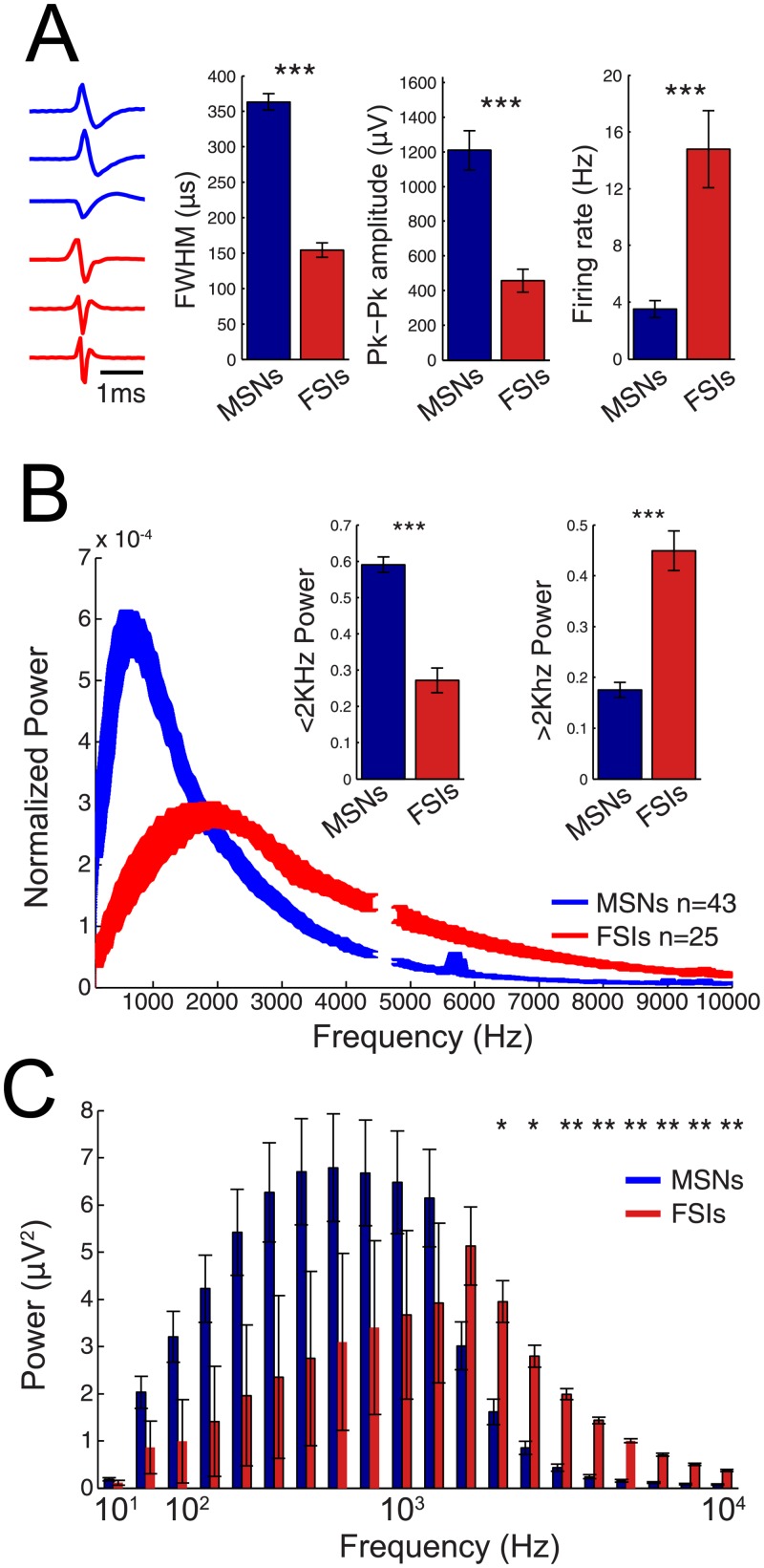
To date, single neuron recordings remain the gold standard for monitoring the activity of neuronal populations. Since obtaining single neuron recordings is not always possible, high frequency or 'multiunit activity' (MUA) is often used as a surrogate. Although MUA recordings allow one to monitor the activity of a large number of neurons, they do not allow identification of specific neuronal subtypes, the knowledge of which is often critical for understanding electrophysiological processes. Here, we explored whether prior knowledge of the single unit waveform of specific neuron types is sufficient to permit the use of MUA to monitor and distinguish differential activity of individual neuron types. We used an experimental and modeling approach to determine if components of the MUA can monitor medium spiny neurons (MSNs) and fast-spiking interneurons (FSIs) in the mouse dorsal striatum. We demonstrate that when well-isolated spikes are recorded, the MUA at frequencies greater than 100Hz is correlated with single unit spiking, highly dependent on the waveform of each neuron type, and accurately reflects the timing and spectral signature of each neuron. However, in the absence of well-isolated spikes (the norm in most MUA recordings), the MUA did not typically contain sufficient information to permit accurate prediction of the respective population activity of MSNs and FSIs. Thus, even under ideal conditions for the MUA to reliably predict the moment-to-moment activity of specific local neuronal ensembles, knowledge of the spike waveform of the underlying neuronal populations is necessary, but not sufficient.
Conflict of interest statement
Figures







Similar articles
-
Representation of the body in the lateral striatum of the freely moving rat: Fast Spiking Interneurons respond to stimulation of individual body parts.Brain Res. 2017 Feb 15;1657:101-108. doi: 10.1016/j.brainres.2016.11.033. Epub 2016 Nov 30. Brain Res. 2017. PMID: 27914882 Free PMC article.
-
Temporal correlations among functionally specialized striatal neural ensembles in reward-conditioned mice.J Neurophysiol. 2016 Mar;115(3):1521-32. doi: 10.1152/jn.01037.2015. Epub 2016 Jan 13. J Neurophysiol. 2016. PMID: 26763779 Free PMC article.
-
Predicting movement from multiunit activity.J Neurosci. 2007 Aug 1;27(31):8387-94. doi: 10.1523/JNEUROSCI.1321-07.2007. J Neurosci. 2007. PMID: 17670985 Free PMC article.
-
Modeling influences of dopamine on synchronization behavior of striatum.Network. 2017;28(1):28-52. doi: 10.1080/0954898X.2017.1378824. Epub 2017 Oct 6. Network. 2017. PMID: 28985088 Review.
-
Striatal function scrutinized through the PAN-TAN-FSI triumvirate.Front Cell Neurosci. 2025 Mar 25;19:1572657. doi: 10.3389/fncel.2025.1572657. eCollection 2025. Front Cell Neurosci. 2025. PMID: 40201383 Free PMC article. Review.
Cited by
-
Novel electrode technologies for neural recordings.Nat Rev Neurosci. 2019 Jun;20(6):330-345. doi: 10.1038/s41583-019-0140-6. Nat Rev Neurosci. 2019. PMID: 30833706 Free PMC article. Review.
-
Differential Sources for 2 Neural Signatures of Target Detection: An Electrocorticography Study.Cereb Cortex. 2018 Jan 1;28(1):9-20. doi: 10.1093/cercor/bhw343. Cereb Cortex. 2018. PMID: 29253249 Free PMC article.
-
Uncovering population contributions to the extracellular potential in the mouse visual system using Laminar Population Analysis.PLoS Comput Biol. 2024 Dec 12;20(12):e1011830. doi: 10.1371/journal.pcbi.1011830. eCollection 2024 Dec. PLoS Comput Biol. 2024. PMID: 39666739 Free PMC article.
-
Uncovering population contributions to the extracellular potential in the mouse visual system using Laminar Population Analysis.bioRxiv [Preprint]. 2024 Jan 16:2024.01.15.575805. doi: 10.1101/2024.01.15.575805. bioRxiv. 2024. Update in: PLoS Comput Biol. 2024 Dec 12;20(12):e1011830. doi: 10.1371/journal.pcbi.1011830. PMID: 38293236 Free PMC article. Updated. Preprint.
-
Evaluation of force feedback in walking using joint torques as "naturalistic" stimuli.J Neurophysiol. 2021 Jul 1;126(1):227-248. doi: 10.1152/jn.00120.2021. Epub 2021 Jun 9. J Neurophysiol. 2021. PMID: 34107221 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
