

Myopathy-inducing mutation H40Y in ACTA1 hampers actin filament structure and function
- PMID: 27112274
- PMCID: PMC4894126
- DOI: 10.1016/j.bbadis.2016.04.013
Myopathy-inducing mutation H40Y in ACTA1 hampers actin filament structure and function
Abstract
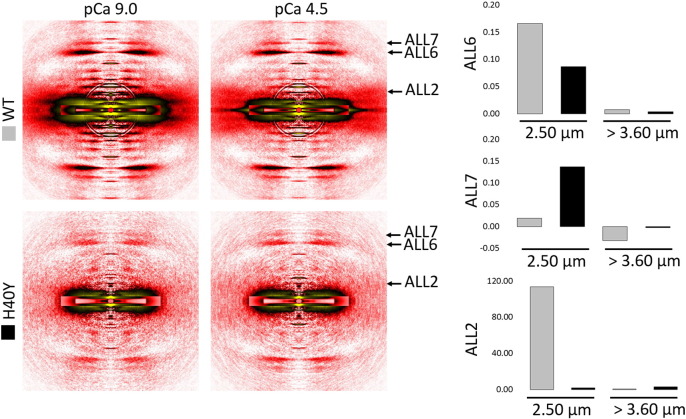
In humans, more than 200 missense mutations have been identified in the ACTA1 gene. The exact molecular mechanisms by which, these particular mutations become toxic and lead to muscle weakness and myopathies remain obscure. To address this, here, we performed a molecular dynamics simulation, and we used a broad range of biophysical assays to determine how the lethal and myopathy-related H40Y amino acid substitution in actin affects the structure, stability, and function of this protein. Interestingly, our results showed that H40Y severely disrupts the DNase I-binding-loop structure and actin filaments. In addition, we observed that normal and mutant actin monomers are likely to form distinctive homopolymers, with mutant filaments being very stiff, and not supporting proper myosin binding. These phenomena underlie the toxicity of H40Y and may be considered as important triggering factors for the contractile dysfunction, muscle weakness and disease phenotype seen in patients.
Keywords: Actin; Contractile dysfunction; In vitro motility assay; Molecular dynamics; Myopathy; Small-angle X-ray scattering.
Copyright © 2016 The Authors. Published by Elsevier B.V. All rights reserved.
Figures



Similar articles
-
Cardiac α-actin over-expression therapy in dominant ACTA1 disease.Hum Mol Genet. 2013 Oct 1;22(19):3987-97. doi: 10.1093/hmg/ddt252. Epub 2013 Jun 4. Hum Mol Genet. 2013. PMID: 23736297
-
Combined MRI and ³¹P-MRS investigations of the ACTA1(H40Y) mouse model of nemaline myopathy show impaired muscle function and altered energy metabolism.PLoS One. 2013 Apr 16;8(4):e61517. doi: 10.1371/journal.pone.0061517. Print 2013. PLoS One. 2013. PMID: 23613869 Free PMC article.
-
ACTA1 H40Y mutant iPSC-derived skeletal myocytes display mitochondrial defects in an in vitro model of nemaline myopathy.Exp Cell Res. 2023 Mar 15;424(2):113507. doi: 10.1016/j.yexcr.2023.113507. Epub 2023 Feb 14. Exp Cell Res. 2023. PMID: 36796746 Free PMC article.
-
Muscle disease caused by mutations in the skeletal muscle alpha-actin gene (ACTA1).Neuromuscul Disord. 2003 Sep;13(7-8):519-31. doi: 10.1016/s0960-8966(03)00101-9. Neuromuscul Disord. 2003. PMID: 12921789 Review.
-
Genotype-phenotype correlations in ACTA1 mutations that cause congenital myopathies.Neuromuscul Disord. 2009 Jan;19(1):6-16. doi: 10.1016/j.nmd.2008.09.005. Epub 2008 Oct 30. Neuromuscul Disord. 2009. PMID: 18976909 Review.
Cited by
-
Nemaline myopathies: a current view.J Muscle Res Cell Motil. 2019 Jun;40(2):111-126. doi: 10.1007/s10974-019-09519-9. Epub 2019 Jun 21. J Muscle Res Cell Motil. 2019. PMID: 31228046 Free PMC article. Review.
-
Identification of Chimeric RNAs in Pig Skeletal Muscle and Transcriptomic Analysis of Chimeric RNA TNNI2-ACTA1 V1.Front Vet Sci. 2021 Oct 27;8:742593. doi: 10.3389/fvets.2021.742593. eCollection 2021. Front Vet Sci. 2021. PMID: 34778431 Free PMC article.
-
Variants in ACTC1 underlie distal arthrogryposis accompanied by congenital heart defects.HGG Adv. 2023 Jun 15;4(3):100213. doi: 10.1016/j.xhgg.2023.100213. eCollection 2023 Jul 13. HGG Adv. 2023. PMID: 37457373 Free PMC article.
-
Nebulin stiffens the thin filament and augments cross-bridge interaction in skeletal muscle.Proc Natl Acad Sci U S A. 2018 Oct 9;115(41):10369-10374. doi: 10.1073/pnas.1804726115. Epub 2018 Sep 24. Proc Natl Acad Sci U S A. 2018. PMID: 30249654 Free PMC article.
-
Pathogenic TNNT1 variants are associated with aberrant thin filament compliance and myofibre hyper-contractility.J Physiol. 2025 Jun;603(12):3533-3550. doi: 10.1113/JP288109. Epub 2025 May 5. J Physiol. 2025. PMID: 40320982 Free PMC article.
References
-
- Nowak K.J., Ravenscroft G., Laing N.G. Skeletal muscle alpha-actin diseases (actinopathies): pathology and mechanisms. Acta Neuropathol. 2013;125:19–32. - PubMed
-
- Nowak K.J., Wattanasirichaigoon D., Goebel H.H., Wilce M., Pelin K., Donner K., Jacob R.L., Hubner C., Oexle K., Anderson J.R., Verity C.M., North K.N., Iannaccone S.T., Muller C.R., Nurnberg P., Muntoni F., Sewry C., Hughes I., Sutphen R., Lacson A.G., Swoboda K.J., Vigneron J., Wallgren-Pettersson C., Beggs A.H., Laing N.G. Mutations in the skeletal muscle alpha-actin gene in patients with actin myopathy and nemaline myopathy. Nat. Genet. 1999;23:208–212. - PubMed
-
- Lehman W., Craig R. Tropomyosin and the steric mechanism of muscle regulation. Adv. Exp. Med. Biol. 2008;644:95–109. - PubMed
-
- Nguyen M.A., Joya J.E., Kee A.J., Domazetovska A., Yang N., Hook J.W., Lemckert F.A., Kettle E., Valova V.A., Robinson P.J., North K.N., Gunning P.W., Mitchell C.A., Hardeman E.C. Hypertrophy and dietary tyrosine ameliorate the phenotypes of a mouse model of severe nemaline myopathy. Brain. 2010 - PubMed
-
- Lindqvist J., Cheng A.J., Renaud G., Hardeman E.C., Ochala J. Distinct underlying mechanisms of limb and respiratory muscle fiber weaknesses in nemaline myopathy. J. Neuropathol. Exp. Neurol. 2013;72:472–481. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
