

CDK1 Prevents Unscheduled PLK4-STIL Complex Assembly in Centriole Biogenesis
- PMID: 27112295
- PMCID: PMC4867225
- DOI: 10.1016/j.cub.2016.03.055
CDK1 Prevents Unscheduled PLK4-STIL Complex Assembly in Centriole Biogenesis
Abstract
Centrioles are essential for the assembly of both centrosomes and cilia. Centriole biogenesis occurs once and only once per cell cycle and is temporally coordinated with cell-cycle progression, ensuring the formation of the right number of centrioles at the right time. The formation of new daughter centrioles is guided by a pre-existing, mother centriole. The proximity between mother and daughter centrioles was proposed to restrict new centriole formation until they separate beyond a critical distance. Paradoxically, mother and daughter centrioles overcome this distance in early mitosis, at a time when triggers for centriole biogenesis Polo-like kinase 4 (PLK4) and its substrate STIL are abundant. Here we show that in mitosis, the mitotic kinase CDK1-CyclinB binds STIL and prevents formation of the PLK4-STIL complex and STIL phosphorylation by PLK4, thus inhibiting untimely onset of centriole biogenesis. After CDK1-CyclinB inactivation upon mitotic exit, PLK4 can bind and phosphorylate STIL in G1, allowing pro-centriole assembly in the subsequent S phase. Our work shows that complementary mechanisms, such as mother-daughter centriole proximity and CDK1-CyclinB interaction with centriolar components, ensure that centriole biogenesis occurs once and only once per cell cycle, raising parallels to the cell-cycle regulation of DNA replication and centromere formation.
Keywords: CDK; PLK4; STIL; centriole duplication; centrosome; licensing; mitosis.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures
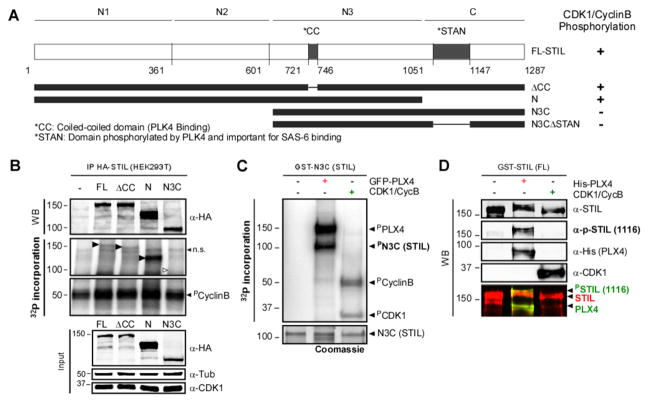
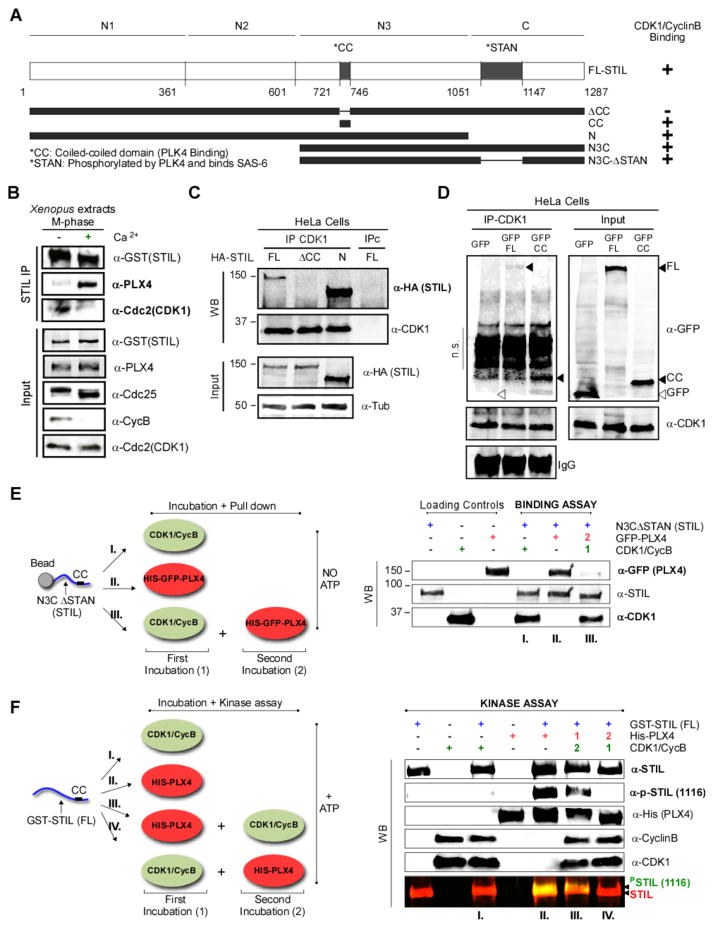
Similar articles
-
The PLK4-STIL-SAS-6 module at the core of centriole duplication.Biochem Soc Trans. 2016 Oct 15;44(5):1253-1263. doi: 10.1042/BST20160116. Biochem Soc Trans. 2016. PMID: 27911707 Free PMC article. Review.
-
PLK4 promotes centriole duplication by phosphorylating STIL to link the procentriole cartwheel to the microtubule wall.Elife. 2019 May 22;8:e46054. doi: 10.7554/eLife.46054. Elife. 2019. PMID: 31115335 Free PMC article.
-
Cell-cycle-regulated expression of STIL controls centriole number in human cells.J Cell Sci. 2012 Mar 1;125(Pt 5):1342-52. doi: 10.1242/jcs.099887. Epub 2012 Feb 20. J Cell Sci. 2012. PMID: 22349698
-
STIL is required for centriole duplication in human cells.J Cell Sci. 2012 Mar 1;125(Pt 5):1353-62. doi: 10.1242/jcs.104109. Epub 2012 Feb 20. J Cell Sci. 2012. PMID: 22349705
-
The centrosome duplication cycle in health and disease.FEBS Lett. 2014 Aug 1;588(15):2366-72. doi: 10.1016/j.febslet.2014.06.030. Epub 2014 Jun 18. FEBS Lett. 2014. PMID: 24951839 Review.
Cited by
-
Regulation of centriolar satellite integrity and its physiology.Cell Mol Life Sci. 2017 Jan;74(2):213-229. doi: 10.1007/s00018-016-2315-x. Epub 2016 Aug 2. Cell Mol Life Sci. 2017. PMID: 27484406 Free PMC article. Review.
-
A key centriole assembly interaction interface between human PLK4 and STIL appears to not be conserved in flies.Biol Open. 2017 Mar 15;6(3):381-389. doi: 10.1242/bio.024661. Biol Open. 2017. PMID: 28202467 Free PMC article.
-
A homeostatic clock sets daughter centriole size in flies.J Cell Biol. 2018 Apr 2;217(4):1233-1248. doi: 10.1083/jcb.201801014. Epub 2018 Mar 2. J Cell Biol. 2018. PMID: 29500190 Free PMC article.
-
Centriole growth is limited by the Cdk/Cyclin-dependent phosphorylation of Ana2/STIL.J Cell Biol. 2022 Sep 5;221(9):e202205058. doi: 10.1083/jcb.202205058. Epub 2022 Jul 21. J Cell Biol. 2022. PMID: 35861803 Free PMC article.
-
Structural and Functional Insights into the Microtubule Organizing Centers of Toxoplasma gondii and Plasmodium spp.Microorganisms. 2021 Dec 3;9(12):2503. doi: 10.3390/microorganisms9122503. Microorganisms. 2021. PMID: 34946106 Free PMC article. Review.
References
-
- Bettencourt-Dias M, Glover DM. Centrosome biogenesis and function: centrosomics brings new understanding. Nat Rev Mol Cell Biol. 2007;8:451–463. - PubMed
-
- Zitouni S, Nabais C, Jana SC, Guerrero A, Bettencourt-Dias M. Polo-like kinases: structural variations lead to multiple functions. Nat Rev Mol Cell Biol. 2014;15:433–452. - PubMed
-
- Bettencourt-Dias M, Rodrigues-Martins A, Carpenter L, Riparbelli M, Lehmann L, Gatt MK, Carmo N, Balloux F, Callaini G, Glover DM. SAK/PLK4 is required for centriole duplication and flagella development. Curr Biol. 2005;15:2199–2207. - PubMed
-
- Habedanck R, Stierhof Y-D, Wilkinson CJ, Nigg EA. The Polo kinase Plk4 functions in centriole duplication. Nat Cell Biol. 2005;7:1140–1146. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
