

Line tension at lipid phase boundaries as driving force for HIV fusion peptide-mediated fusion
- PMID: 27113279
- PMCID: PMC4853434
- DOI: 10.1038/ncomms11401
Line tension at lipid phase boundaries as driving force for HIV fusion peptide-mediated fusion
Abstract
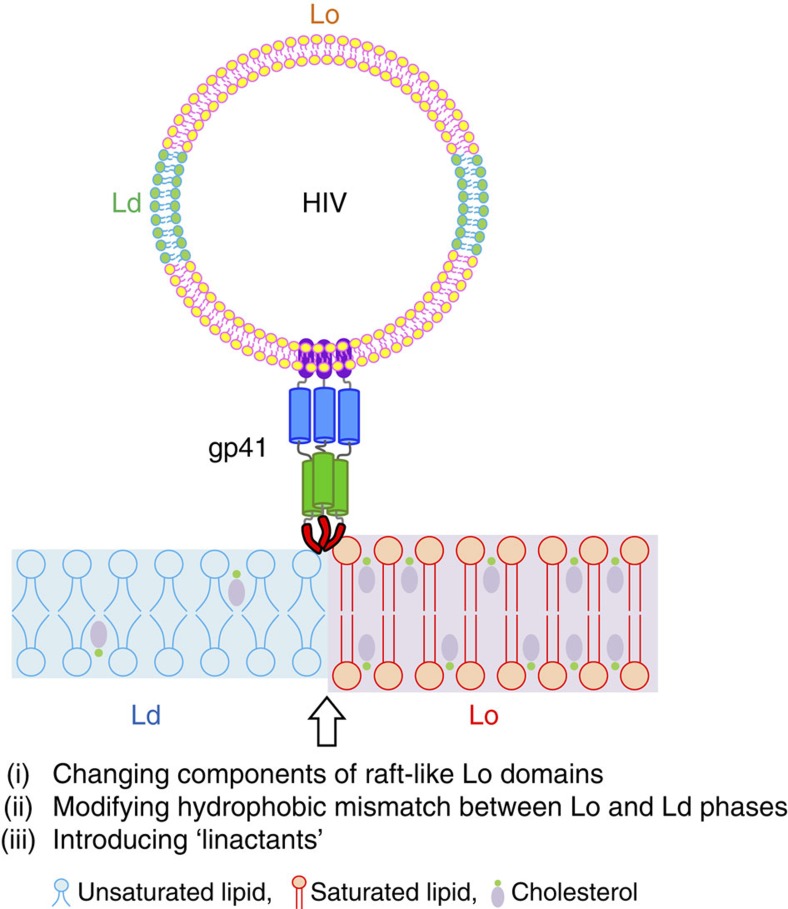
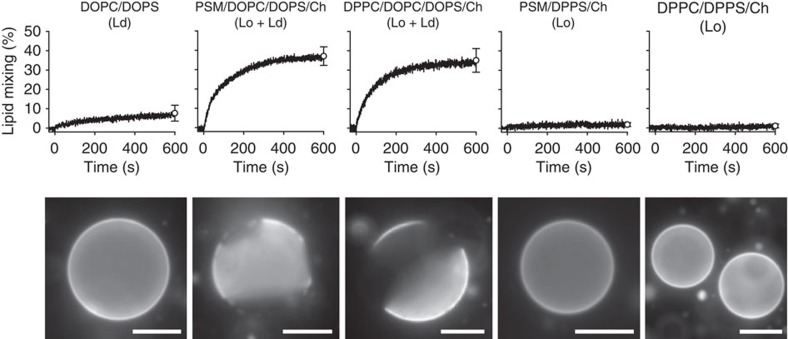
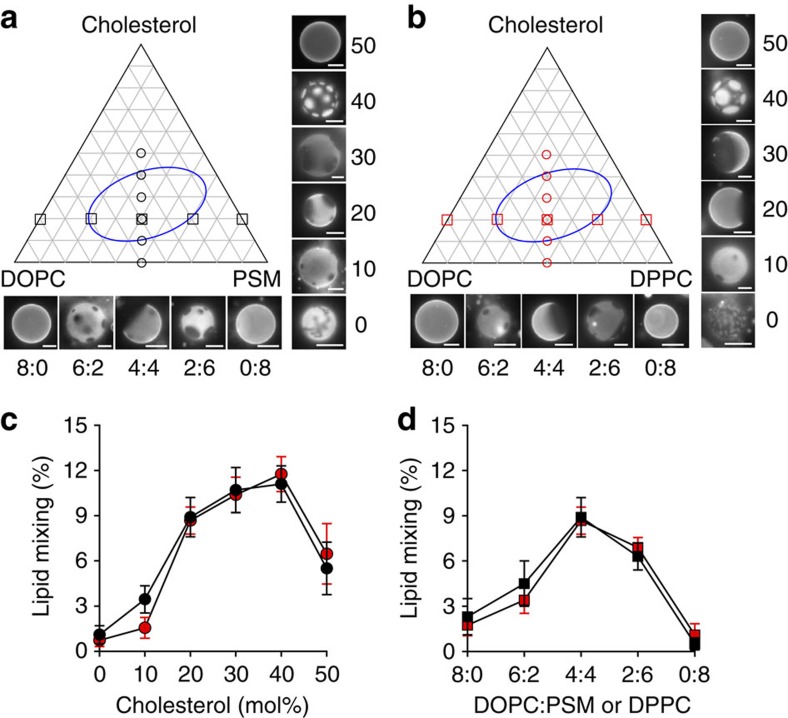
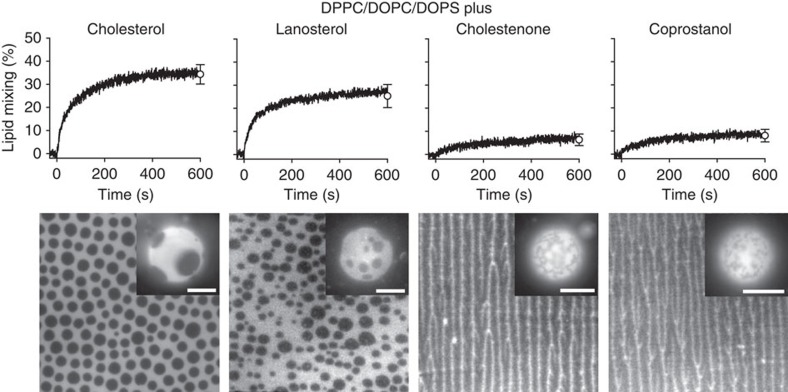
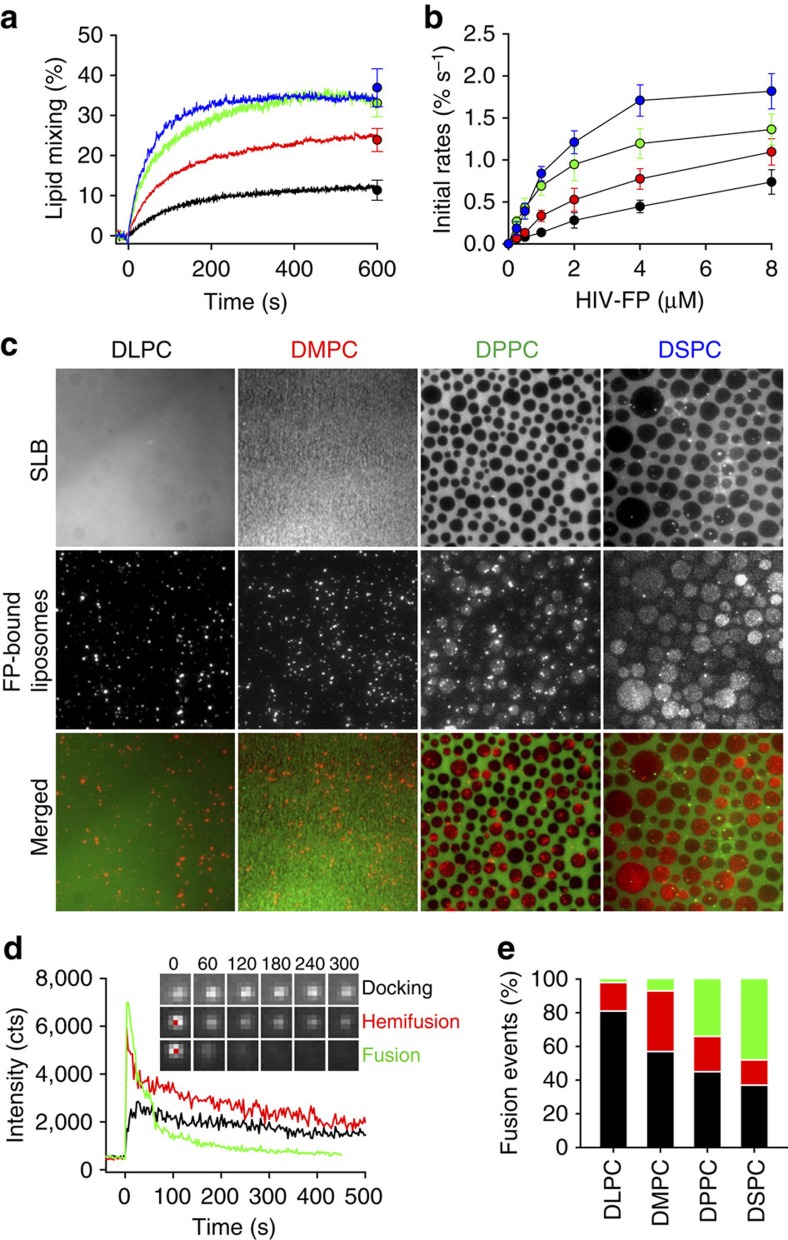
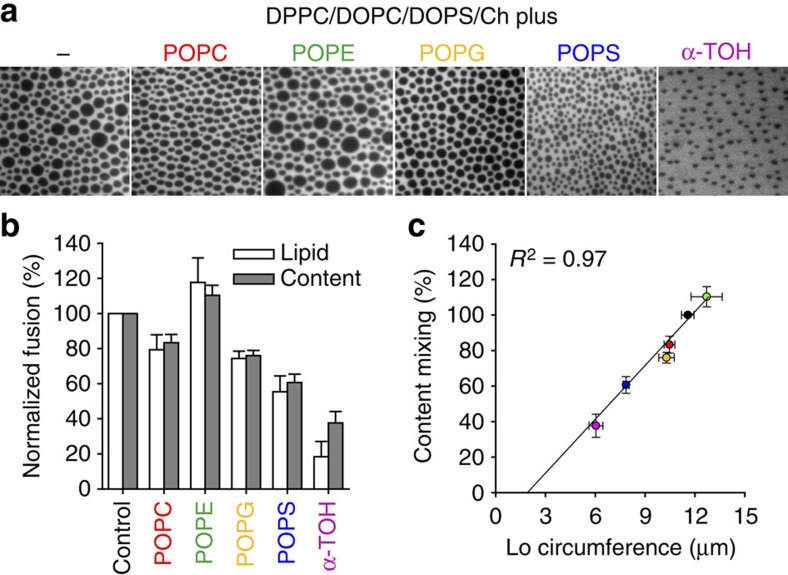
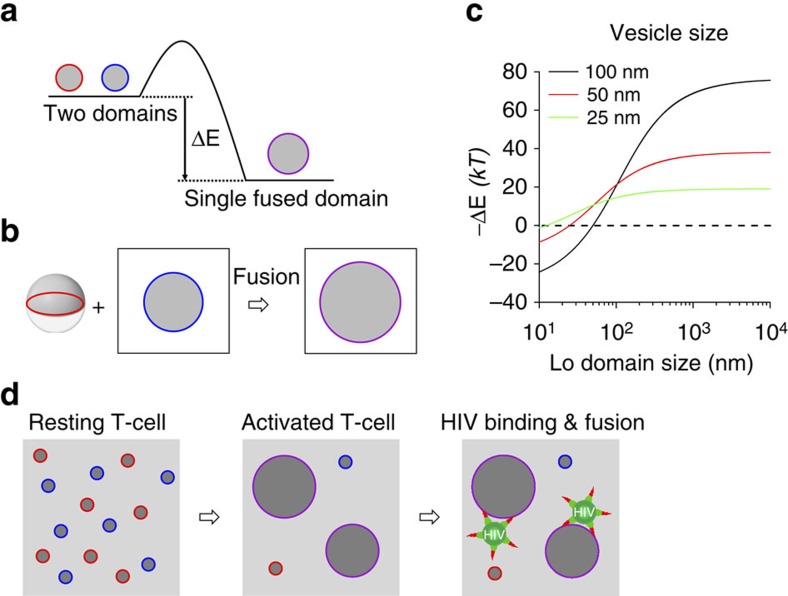
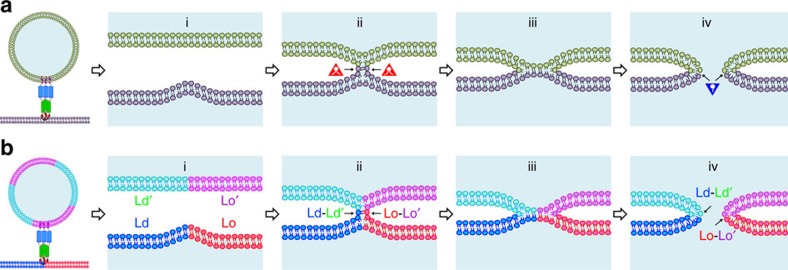
Lipids and proteins are organized in cellular membranes in clusters, often called 'lipid rafts'. Although raft-constituent ordered lipid domains are thought to be energetically unfavourable for membrane fusion, rafts have long been implicated in many biological fusion processes. For the case of HIV gp41-mediated membrane fusion, this apparent contradiction can be resolved by recognizing that the interfaces between ordered and disordered lipid domains are the predominant sites of fusion. Here we show that line tension at lipid domain boundaries contributes significant energy to drive gp41-fusion peptide-mediated fusion. This energy, which depends on the hydrophobic mismatch between ordered and disordered lipid domains, may contribute tens of kBT to fusion, that is, it is comparable to the energy required to form a lipid stalk intermediate. Line-active compounds such as vitamin E lower line tension in inhomogeneous membranes, thereby inhibit membrane fusion, and thus may be useful natural viral entry inhibitors.
Figures
References
-
- Tamm L. K. & Han X. Viral fusion peptides: a tool set to disrupt and connect biological membranes. Biosci. Rep. 20, 501–518 (2000). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
