

Ghosts in the Machine II: Neural Correlates of Memory Interference from the Previous Trial
- PMID: 27114176
- PMCID: PMC6059123
- DOI: 10.1093/cercor/bhw106
Ghosts in the Machine II: Neural Correlates of Memory Interference from the Previous Trial
Abstract
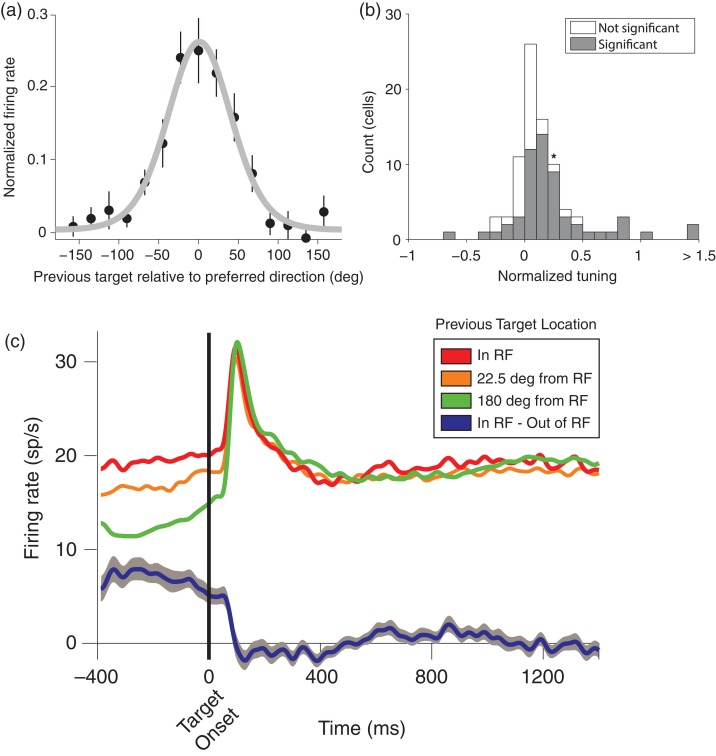
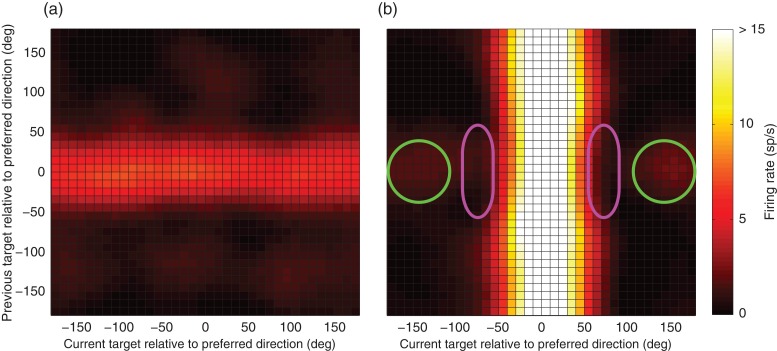
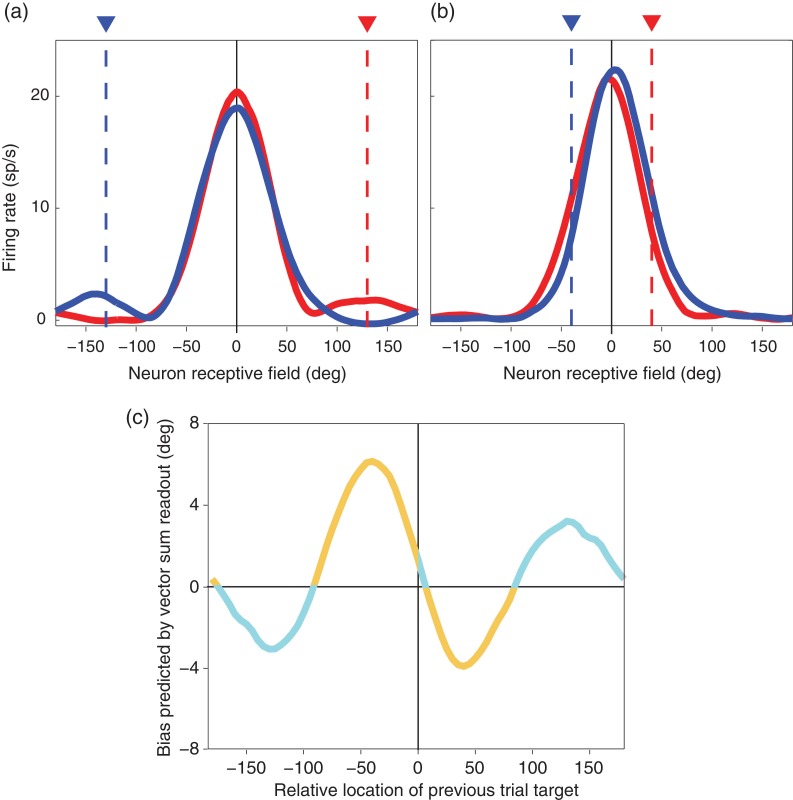
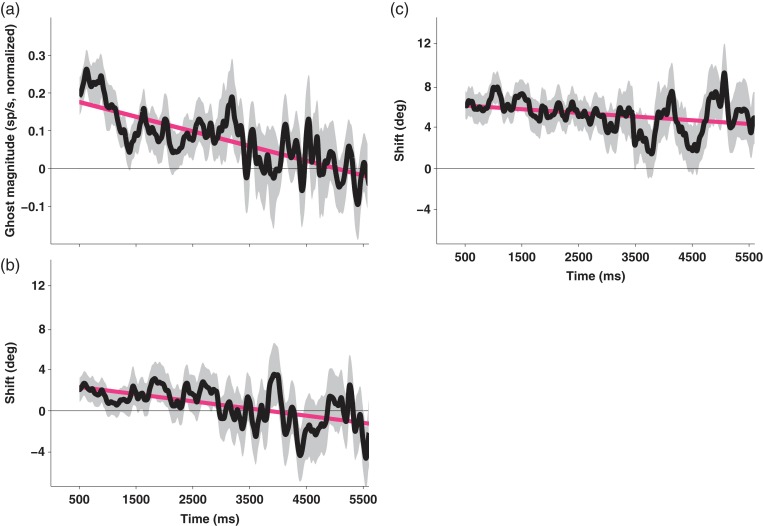
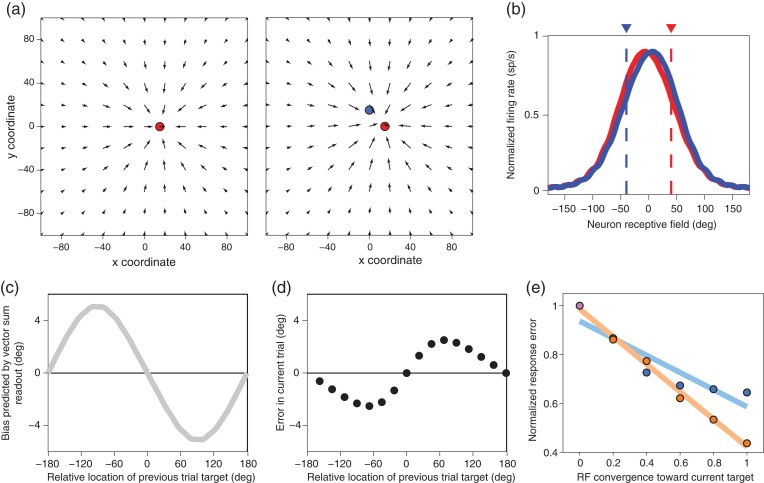
Previous memoranda interfere with working memory. For example, spatial memories are biased toward locations memorized on the previous trial. We predicted, based on attractor network models of memory, that activity in the frontal eye fields (FEFs) encoding a previous target location can persist into the subsequent trial and that this ghost will then bias the readout of the current target. Contrary to this prediction, we find that FEF memory representations appear biased away from (not toward) the previous target location. The behavioral and neural data can be reconciled by a model in which receptive fields of memory neurons converge toward remembered locations, much as receptive fields converge toward attended locations. Convergence increases the resources available to encode the relevant memoranda and decreases overall error in the network, but the residual convergence from the previous trial can give rise to an attractive behavioral bias on the next trial.
Keywords: attractor network models; frontal eye fields; proactive interference; receptive field remapping; spatial working memory.
© The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please e-mail: journals.permissions@oup.com.
Figures



References
-
- Abbott LF. 1994. Decoding neuronal firing and modelling neural networks. Q Rev Biophys. 27:291–331. - PubMed
-
- Bruce CJ, Goldberg ME. 1985. Primate frontal eye fields. I. Single neurons discharging before saccades. J Neurophysiol. 53:603–635. - PubMed
-
- Bruce CJ, Goldberg ME, Bushnell MC, Stanton GB. 1985. Primate frontal eye fields. II. Physiological and anatomical correlates of electrically evoked eye movements. J Neurophysiol. 54:714–734. - PubMed
-
- Chafee MV, Goldman-Rakic PS. 1998. Matching patterns of activity in primate prefrontal area 8a and parietal area 7ip neurons during a spatial working memory task. J Neurophysiol. 79:2919–2940. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
